From Surf Wiki (app.surf) — the open knowledge base
Ubiquitin ligase
Protein
Protein
| Field | Value |
|---|---|
| Name | Ubiquitin—protein ligase |
| EC_number | 2.3.2.27 |
| CAS_number | 74812-49-0 |
| GO_code | 0061630 |
| image | 4a4c.png |
| caption | E3 ubiquitin ligase Cbl (blue) in complex with E2 (cyan) and substrate peptide (green). PDB entry |
A ubiquitin ligase (also called an E3 ubiquitin ligase) is a protein that recruits an E2 ubiquitin-conjugating enzyme that has been loaded with ubiquitin, recognizes a protein substrate, and assists or directly catalyzes the transfer of ubiquitin from the E2 to the protein substrate. In simple and more general terms, the ligase enables movement of ubiquitin from a ubiquitin carrier to another protein (the substrate) by some mechanism. The ubiquitin, once it reaches its destination, ends up being attached by an isopeptide bond to a lysine residue, which is part of the target protein. E3 ligases interact with both the target protein and the E2 enzyme, and so impart substrate specificity to the E2. Commonly, E3s polyubiquitinate their substrate with Lys48-linked chains of ubiquitin, targeting the substrate for destruction by the proteasome. However, many other types of linkages are possible and alter a protein's activity, interactions, or localization. Ubiquitination by E3 ligases regulates diverse areas such as cell trafficking, DNA repair, and signaling and is of profound importance in cell biology. E3 ligases are also key players in cell cycle control, mediating the degradation of cyclins, as well as cyclin dependent kinase inhibitor proteins. The human genome encodes over 600 putative E3 ligases, allowing for tremendous diversity in substrates. Certain E3 ligases have been utilized in targeted protein degradation applications.
Ubiquitination system
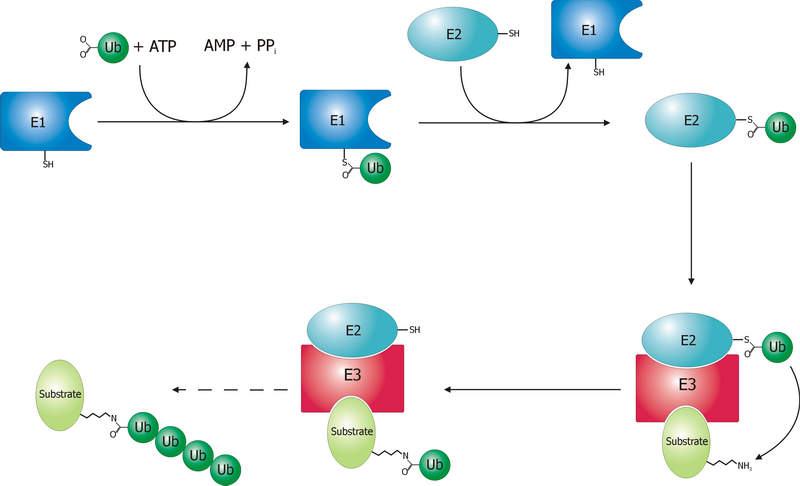
The ubiquitin ligase is referred to as an E3, and operates in conjunction with an E1 ubiquitin-activating enzyme and an E2 ubiquitin-conjugating enzyme. There is one major E1 enzyme, shared by all ubiquitin ligases, that uses ATP to activate ubiquitin for conjugation and transfers it to an E2 enzyme. The E2 enzyme interacts with a specific E3 partner and transfers the ubiquitin to the target protein. The E3, which may be a multi-protein complex, is, in general, responsible for targeting ubiquitination to specific substrate proteins.
The ubiquitylation reaction proceeds in three or four steps depending on the mechanism of action of the E3 ubiquitin ligase. In the conserved first step, an E1 cysteine residue attacks the ATP-activated C-terminal glycine on ubiquitin, resulting in a thioester Ub-S-E1 complex. The energy from ATP and diphosphate hydrolysis drives the formation of this reactive thioester, and subsequent steps are thermoneutral. Next, a transthiolation reaction occurs, in which an E2 cysteine residue attacks and replaces the E1. HECT domain type E3 ligases will have one more transthiolation reaction to transfer the ubiquitin molecule onto the E3, whereas the much more common RING finger domain type ligases transfer ubiquitin directly from E2 to the substrate. The final step in the first ubiquitylation event is an attack from the target protein lysine amine group, which will remove the cysteine, and form a stable isopeptide bond. One notable exception to this is p21 protein, which appears to be ubiquitylated using its N-terminal amine, thus forming a peptide bond with ubiquitin.
Ubiquitin ligase families
Humans have an estimated 500-1000 E3 ligases, which impart substrate specificity onto the E1 and E2. The E3 ligases are classified into four families: HECT, RING-finger, U-box, and PHD-finger. The RING-finger E3 ligases are the largest family and contain ligases such as the anaphase-promoting complex (APC) and the SCF complex (Skp1-Cullin-F-box protein complex). SCF complexes consist of four proteins: Rbx1, Cul1, Skp1, which are invariant among SCF complexes, and an F-box protein, which varies. Around 70 human F-box proteins have been identified. F-box proteins contain an F-box, which binds the rest of the SCF complex, and a substrate binding domain, which gives the E3 its substrate specificity.
Mono- and poly-ubiquitylation

Ubiquitin signaling relies on the diversity of ubiquitin tags for the specificity of its message. A protein can be tagged with a single ubiquitin molecule (monoubiquitylation), or variety of different chains of ubiquitin molecules (polyubiquitylation). E3 ubiquitin ligases catalyze polyubiquitination events much in the same way as the single ubiquitylation mechanism, using instead a lysine residue from a ubiquitin molecule currently attached to substrate protein to attack the C-terminus of a new ubiquitin molecule. For example, a common 4-ubiquitin tag, linked through the lysine at position 48 (K48) recruits the tagged protein to the proteasome, and subsequent degradation. However, all seven of the ubiquitin lysine residues (K6, K11, K27, K29, K33, K48, and K63), as well as the N-terminal methionine are used in chains in vivo.
Monoubiquitination has been linked to membrane protein endocytosis pathways. For example, phosphorylation of the Tyrosine at position 1045 in the Epidermal Growth Factor Receptor (EGFR) can recruit the RING type E3 ligase c-Cbl, via an SH2 domain. C-Cbl monoubiquitylates EGFR, signaling for its internalization and trafficking to the lysosome.
Monoubiquitination also can regulate cytosolic protein localization. For example, the E3 ligase MDM2 ubiquitylates p53 either for degradation (K48 polyubiquitin chain), or for nuclear export (monoubiquitylation). These events occur in a concentration dependent fashion, suggesting that modulating E3 ligase concentration is a cellular regulatory strategy for controlling protein homeostasis and localization.
Substrate recognition
Ubiquitin ligases are the final, and potentially the most important determinant of substrate specificity in ubiquitination of proteins. The ligases must simultaneously distinguish their protein substrate from thousands of other proteins in the cell, and from other (ubiquitination-inactive) forms of the same protein. This can be achieved by different mechanisms, most of which involve recognition of degrons: specific short amino acid sequences or chemical motifs on the substrate.
N-degrons
Proteolytic cleavage can lead to exposure of residues at the N-terminus of a protein. According to the N-end rule, different N-terminal amino acids (or N-degrons) are recognized to a different extent by their appropriate ubiquitin ligase (N-recognin), influencing the half-life of the protein. For instance, positively charged (Arg, Lys, His) and bulky hydrophobic amino acids (Phe, Trp, Tyr, Leu, Ile) are recognized preferentially and thus considered destabilizing degrons since they allow faster degradation of their proteins.
Phosphodegrons

A degron can be converted into its active form by a post-translational modification such as phosphorylation of a tyrosine, serine or threonine residue. In this case, the ubiquitin ligase exclusively recognizes the phosphorylated version of the substrate due to stabilization within the binding site. For example, FBW7, the F-box substrate recognition unit of an SCFFBW7ubiquitin ligase, stabilizes a phosphorylated substrate by hydrogen binding its arginine residues to the phosphate, as shown in the figure to the right. In absence of the phosphate, residues of FBW7 repel the substrate.
Oxygen and small molecule dependent degrons
The presence of oxygen or other small molecules can influence degron recognition. The von Hippel-Lindau (VHL) protein (substrate recognition part of a specific E3 ligase), for instance, recognizes the hypoxia-inducible factor alpha (HIF-α) only under normal oxygen conditions, when its proline is hydroxylated. Under hypoxia, on the other hand, HIF-a is not hydroxylated, evades ubiquitination and thus operates in the cell at higher concentrations which can initiate transcriptional response to hypoxia. Another example of small molecule control of protein degradation is phytohormone auxin in plants. Auxin binds to TIR1 (the substrate recognition domain of SCFTIR1ubiquitin ligase) increasing the affinity of TIR1 for its substrates (transcriptional repressors: Aux/IAA), and promoting their degradation.
Misfolded and sugar degrons
In addition to recognizing amino acids, ubiquitin ligases can also detect unusual features on substrates that serve as signals for their destruction. For example, San1 (Sir antagonist 1), a nuclear protein quality control in yeast, has a disordered substrate binding domain, which allows it to bind to hydrophobic domains of misfolded proteins. Misfolded or excess unassembled glycoproteins of the ERAD pathway, on the other hand, are recognized by Fbs1 and Fbs2, mammalian F-box proteins of E3 ligases SCFFbs1and SCFFbs2. These recognition domains have small hydrophobic pockets allowing them to bind high-mannose containing glycans.
Structural motifs
In addition to linear degrons, the E3 ligase can in some cases also recognize structural motifs on the substrate. In this case, the 3D motif can allow the substrate to directly relate its biochemical function to ubiquitination. This relation can be demonstrated with TRF1 protein (regulator of human telomere length), which is recognized by its corresponding E3 ligase (FBXO4) via an intermolecular beta sheet interaction. TRF1 cannot be ubiquinated while telomere bound, likely because the same TRF1 domain that binds to its E3 ligase also binds to telomeres.
Disease relevance
E3 ubiquitin ligases regulate homeostasis, cell cycle, and DNA repair pathways, and as a result, a number of these proteins are involved in a variety of cancers, including famously MDM2, BRCA1, and Von Hippel-Lindau tumor suppressor. For example, a mutation of MDM2 has been found in stomach cancer, renal cell carcinoma, and liver cancer (amongst others) to deregulate MDM2 concentrations by increasing its promoter's affinity for the Sp1 transcription factor, causing increased transcription of MDM2 mRNA. such as proximity-dependent biotin identification (BioID), ubiquitin ligase-substrate trapping, and tandem ubiquitin-binding entities (TUBEs).
Examples
- A RING (Really Interesting New Gene) domain binds the E2 conjugase and might be found to mediate enzymatic activity in the E2-E3 complex
- An F-box domain (as in the SCF complex) binds the ubiquitinated substrate. (e.g., Cdc 4, which binds the target protein Sic1; Grr1, which binds Cln).
- A HECT domain, which is involved in the transfer of ubiquitin from the E2 to the substrate.
Targeted protein degradation
In 2001, work from the labs of Craig Crews and Raymond Deshaies described the development of proteolysis-targeting chimeras (PROTACs). Using a small molecule to recruit an E3 ubiquitin ligase to a target protein, this work demonstrated that induced proximity could be used to effect the ubiquitination and proteasomal degradation of a target protein. PROTACs have been frequently applied using the E3 ubiquitin ligases CRBN and VHL to degrade various targets of biological and therapeutic relevance. Multiple groups have sought out additional E3 ligases to co-opt for targeted protein degradation such as FBXO22 and KLHDC2.
While PROTACs generally are heterobifunctional compounds linking an E3 ligase binder to a target protein binder, molecular glues also exist that induce protein-protein interactions with E3 ligases, leading to degradation of various substrate proteins. Molecular glues often have been discovered through serendipity, though various methodologies have been explored to expedite the discovery of molecular glues.
Biologic modalities for targeted protein degradation have also been explored by fusing E3 ligases to target recognition domains such as nanobodies. These modalities are sometimes referred to as bioPROTACs. While bioPROTACs are advantageous for targeting proteins lacking small molecule ligands, challenges in delivery, pharmacokinetics, and immunogenicity have so far precluded clinical development. Studies exploring different delivery mechanisms have sought to address these shortcomings. In another variant of this idea, bispecific antibodies to recruit membrane-bound E3 ligases to cell surface proteins (AbTACs) have also been developed.
Individual E3 ubiquitin ligases
- E3A
- mdm2
- Anaphase-promoting complex (APC)
- UBR5 (EDD1)
- SOCS/ BC-box/ eloBC/ CUL5/ RING
- LNXp80
- CBX4, CBLL1
- HACE1
- HECTD1, HECTD2, HECTD3, HECTD4
- HECW1, HECW2
- HERC1, HERC2, HERC3, HERC4, HERC5, HERC6
- HUWE1, ITCH
- NEDD4, NEDD4L
- PPIL2
- PRPF19
- PIAS1, PIAS2, PIAS3, PIAS4
- RANBP2
- RNF4, RNF167
- RBX1
- SMURF1, SMURF2
- STUB1
- TOPORS
- TRIP12
- UBE3A, UBE3B, UBE3C, UBE3D
- UBE4A, UBE4B
- UBOX5
- UBR5
- VHL
- WWP1, WWP2
- Parkin
- MKRN1
References
References
- (January 2012). "Structural basis for autoinhibition and phosphorylation-dependent activation of c-Cbl". Nature Structural & Molecular Biology.
- (1998). "The ubiquitin system". Annual Review of Biochemistry.
- (2013). "Ubiquitin ligases and cell cycle control". Annual Review of Biochemistry.
- (January 2008). "Genome-wide and functional annotation of human E3 ubiquitin ligases identifies MULAN, a mitochondrial E3 that regulates the organelle's dynamics and signaling". PLOS ONE.
- (March 2022). "PROTAC targeted protein degraders: the past is prologue". Nature Reviews Drug Discovery.
- (February 2012). "HECT and RING finger families of E3 ubiquitin ligases at a glance". Journal of Cell Science.
- Walsh, Christopher. (March 2015). "Posttranslational Modification of Proteins: Expanding Nature's Inventory". Roberts.
- (October 2003). "Proteasome-mediated degradation of p21 via N-terminal ubiquitinylation". Cell.
- (May 2006). "Ubiquitin ligases: cell-cycle control and cancer". Nature Reviews. Cancer.
- (November 2004). "Systematic analysis and nomenclature of mammalian F-box proteins". Genes & Development.
- (April 1987). "Structure of ubiquitin refined at 1.8 A resolution". Journal of Molecular Biology.
- (May 2011). "Constructing and decoding unconventional ubiquitin chains". Nature Structural & Molecular Biology.
- (2003). "Signals for sorting of transmembrane proteins to endosomes and lysosomes". Annual Review of Biochemistry.
- (December 2003). "Mono- versus polyubiquitination: differential control of p53 fate by Mdm2". Science.
- (June 2017). "Ubiquitin Ligases: Structure, Function, and Regulation". Annual Review of Biochemistry.
- (September 2008). "Diversity of degradation signals in the ubiquitin-proteasome system". Nature Reviews Molecular Cell Biology.
- (October 2011). "The N-end rule pathway: emerging functions and molecular principles of substrate recognition". Nature Reviews Molecular Cell Biology.
- (2012). "The N-end rule pathway". Annual Review of Biochemistry.
- (June 2017). "Recognition of substrate degrons by E3 ubiquitin ligases and modulation by small-molecule mimicry strategies". Current Opinion in Structural Biology.
- (September 2015). "Expanding the ubiquitin code through post-translational modification". EMBO Reports.
- (September 2013). "Phospho-Ser/Thr-binding domains: navigating the cell cycle and DNA damage response". Nature Reviews Molecular Cell Biology.
- (April 2001). "Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation". Science.
- (April 2014). "Plant ubiquitin ligases as signaling hubs". Nature Structural & Molecular Biology.
- (2019-02-19). "Sugar-Recognizing Ubiquitin Ligases: Action Mechanisms and Physiology". Frontiers in Physiology.
- (August 2011). "RINGs of good and evil: RING finger ubiquitin ligases at the crossroads of tumour suppression and oncogenesis". Nature Reviews. Cancer.
- (January 2015). "Role of E3 ubiquitin ligases in gastric cancer". World Journal of Gastroenterology.
- (February 2015). "The T309G murine double minute 2 gene polymorphism is an independent prognostic factor for patients with renal cell carcinoma". DNA and Cell Biology.
- (November 2014). "Association between murine double minute 2 T309G polymorphism and risk of liver cancer". Tumour Biology.
- (July 2019). "Using proteomics to identify ubiquitin ligase-substrate pairs: how novel methods may unveil therapeutic targets for neurodegenerative diseases". Cellular and Molecular Life Sciences.
- (2005). "E3 ubiquitin ligases". Essays in Biochemistry.
- (July 1996). "SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box". Cell.
- (2001-07-17). "Protacs: chimeric molecules that target proteins to the Skp1-Cullin-F box complex for ubiquitination and degradation". Proceedings of the National Academy of Sciences of the United States of America.
- (2015-06-18). "Hijacking the E3 Ubiquitin Ligase Cereblon to Efficiently Target BRD4". Chemistry & Biology.
- (2015-06-19). "Phthalimide conjugation as a strategy for in vivo target protein degradation". Science.
- (August 2015). "Catalytic in vivo protein knockdown by small-molecule PROTACs". Nature Chemical Biology.
- (2015-08-21). "Selective Small Molecule Induced Degradation of the BET Bromodomain Protein BRD4". ACS Chemical Biology.
- (December 2024). "Recruitment of FBXO22 for targeted degradation of NSD2". Nature Chemical Biology.
- (December 2024). "A CRISPR activation screen identifies FBXO22 supporting targeted protein degradation". Nature Chemical Biology.
- (2024-06-26). "Alkylamine-tethered molecules recruit FBXO22 for targeted protein degradation". Nature Communications.
- (2024-10-12). "Principles of paralog-specific targeted protein degradation engaging the C-degron E3 KLHDC2". Nature Communications.
- (2010-03-12). "Identification of a Primary Target of Thalidomide Teratogenicity". Science.
- (1996-07-12). "Structure of the FKBP12-rapamycin complex interacting with the binding domain of human FRAP". Science.
- (1991-08-23). "Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes". Cell.
- (2025-03-19). "Discovery of a VHL molecular glue degrader of GEMIN3 by Picowell RNA-seq".
- (2024-11-20). "Discovery of CRBN-Dependent WEE1 Molecular Glue Degraders from a Multicomponent Combinatorial Library". Journal of the American Chemical Society.
- (2024-09-26). "Charged Molecular Glue Discovery Enabled by Targeted Degron Display".
- (2023-12-19). "Direct-to-biology, automated, nano-scale synthesis, and phenotypic screening-enabled E3 ligase modulator discovery". Nature Communications.
- (2023-05-24). "Rational Chemical Design of Molecular Glue Degraders". ACS Central Science.
- (2024-10-04). "Unveiling the hidden interactome of CRBN molecular glues with chemoproteomics".
- (2020-03-17). "bioPROTACs as versatile modulators of intracellular therapeutic targets including proliferating cell nuclear antigen (PCNA)". Proceedings of the National Academy of Sciences.
- (2023-11-04). "A TRIM21-based bioPROTAC highlights the therapeutic benefit of HuR degradation". Nature Communications.
- (2025-02-26). "Protein-Based Degraders: From Chemical Biology Tools to Neo-Therapeutics". Chemical Reviews.
- (2024-07-10). "Lipid-mediated intracellular delivery of recombinant bioPROTACs for the rapid degradation of undruggable proteins". Nature Communications.
- (2021-01-20). "Development of Antibody-Based PROTACs for the Degradation of the Cell-Surface Immune Checkpoint Protein PD-L1". Journal of the American Chemical Society.
This article was imported from Wikipedia and is available under the Creative Commons Attribution-ShareAlike 4.0 License. Content has been adapted to SurfDoc format. Original contributors can be found on the article history page.
Ask Mako anything about Ubiquitin ligase — get instant answers, deeper analysis, and related topics.
Research with MakoFree with your Surf account
Create a free account to save articles, ask Mako questions, and organize your research.
Sign up freeThis content may have been generated or modified by AI. CloudSurf Software LLC is not responsible for the accuracy, completeness, or reliability of AI-generated content. Always verify important information from primary sources.
Report