From Surf Wiki (app.surf) — the open knowledge base
Oxidative phosphorylation
Metabolic pathway

Metabolic pathway

Oxidative phosphorylation(UK , US : or electron transport-linked phosphorylation or terminal oxidation, is the metabolic pathway in which cells use enzymes to oxidize nutrients, thereby releasing chemical energy in order to produce adenosine triphosphate (ATP). In eukaryotes, this takes place inside mitochondria. Almost all aerobic organisms carry out oxidative phosphorylation. This pathway is so pervasive because it releases more energy than fermentation.
In aerobic respiration, the energy stored in the chemical bonds of glucose is released by the cell in glycolysis and subsequently the citric acid cycle, producing carbon dioxide and the energetic electron donors NADH and FADH₂. Oxidative phosphorylation uses these molecules and O2 to produce ATP, which is used throughout the cell whenever energy is needed. During oxidative phosphorylation, electrons are transferred from the electron donors to a series of electron acceptors in a series of redox reactions ending in oxygen, whose reaction releases half of the total energy.
In eukaryotes, these redox reactions are catalyzed by a series of protein complexes within the inner mitochondrial membrane; whereas, in prokaryotes, these proteins are located in the cell's plasma membrane. These linked sets of proteins are called the electron transport chain. In mitochondria, five main protein complexes are involved, whereas prokaryotes have various other enzymes, using a variety of electron donors and acceptors.
The energy transferred by electrons flowing through this electron transport chain is used to transport protons across the inner membrane. This generates potential energy in the form of a pH gradient and the resulting electrical potential across this membrane. This store of energy is tapped when protons flow back across the membrane through ATP synthase in a process called chemiosmosis. The ATP synthase uses the energy to transform adenosine diphosphate (ADP) into adenosine triphosphate, in a phosphorylation reaction. The reaction is driven by the proton flow, which forces the rotation of a part of the enzyme. The ATP synthase is a rotary mechanical motor.
Although oxidative phosphorylation is a vital part of metabolism, it produces reactive oxygen species such as superoxide and hydrogen peroxide, which lead to propagation of free radicals, damaging cells and contributing to disease and, possibly, aging and senescence. The enzymes carrying out this metabolic pathway are also the target of many drugs and poisons that inhibit their activities.
Chemiosmosis
Oxidative phosphorylation works by using energy-releasing chemical reactions to drive energy-requiring reactions. The two sets of reactions are said to be coupled. This means one cannot occur without the other. The chain of redox reactions driving the flow of electrons through the electron transport chain, from electron donors such as NADH to electron acceptors such as oxygen and hydrogen (protons), is an exergonic process – it releases energy, whereas the synthesis of ATP is an endergonic process, which requires an input of energy. Both the electron transport chain and the ATP synthase are embedded in a membrane, and energy is transferred from the electron transport chain to the ATP synthase by movements of protons across this membrane, in a process called chemiosmosis. A current of protons is driven from the negative N-side of the membrane to the positive P-side through the proton-pumping enzymes of the electron transport chain. The movement of protons creates an electrochemical gradient across the membrane, which is called the proton-motive force. It has two components: a difference in proton concentration (a H+ gradient, ΔpH) and a difference in electric potential, with the N-side having a negative charge.
ATP synthase releases this stored energy by completing the circuit and allowing protons to flow down the electrochemical gradient, back to the N-side of the membrane. The electrochemical gradient drives the rotation of part of the enzyme's structure and couples this motion to the synthesis of ATP.
The two components of the proton-motive force are thermodynamically equivalent: In mitochondria, the largest part of energy is provided by the potential; in alkaliphile bacteria the electrical energy even has to compensate for a counteracting inverse pH difference. Inversely, chloroplasts operate mainly on ΔpH. However, they also require a small membrane potential for the kinetics of ATP synthesis. In the case of the fusobacterium Propionigenium modestum it drives the counter-rotation of subunits a and c of the FO motor of ATP synthase.
The amount of energy released by oxidative phosphorylation is high, compared with the amount produced by fermentation. Glycolysis produces only 2 ATP molecules, but somewhere between 30 and 36 ATPs are produced by the oxidative phosphorylation of the 10 NADH and 2 succinate molecules made by converting one molecule of glucose to carbon dioxide and water, while each cycle of beta oxidation of a fatty acid yields about 14 ATPs. These ATP yields are theoretical maximum values; in practice, some protons leak across the membrane, lowering the yield of ATP.
Electron and proton transfer molecules

The electron transport chain carries both protons and electrons, passing electrons from donors to acceptors, and transporting protons across a membrane. These processes use both soluble and protein-bound transfer molecules. In the mitochondria, electrons are transferred within the intermembrane space by the water-soluble electron transfer protein cytochrome c. This carries only electrons, and these are transferred by the reduction and oxidation of an iron atom that the protein holds within a heme group in its structure. Cytochrome c is also found in some bacteria, where it is located within the periplasmic space.
Within the inner mitochondrial membrane, the lipid-soluble electron carrier coenzyme Q10 (Q) carries both electrons and protons by a redox cycle. This small benzoquinone molecule is very hydrophobic, so it diffuses freely within the membrane. When Q accepts two electrons and two protons, it becomes reduced to the ubiquinol form (QH2); when QH2 releases two electrons and two protons, it becomes oxidized back to the ubiquinone (Q) form. As a result, if two enzymes are arranged so that Q is reduced on one side of the membrane and QH2 oxidized on the other, ubiquinone will couple these reactions and shuttle protons across the membrane. Some bacterial electron transport chains use different quinones, such as menaquinone, in addition to ubiquinone.
Within proteins, electrons are transferred between flavin cofactors, iron–sulfur clusters and cytochromes. There are several types of iron–sulfur cluster. The simplest kind found in the electron transfer chain consists of two iron atoms joined by two atoms of inorganic sulfur; these are called [2Fe–2S] clusters. The second kind, called [4Fe–4S], contains a cube of four iron atoms and four sulfur atoms. Each iron atom in these clusters is coordinated by an additional amino acid, usually by the sulfur atom of cysteine. Metal ion cofactors undergo redox reactions without binding or releasing protons, so in the electron transport chain they serve solely to transport electrons through proteins. Electrons move quite long distances through proteins by hopping along chains of these cofactors. This occurs by quantum tunnelling, which is rapid over distances of less than 1.4 m.
Eukaryotic electron transport chains
Many catabolic biochemical processes, such as glycolysis, the citric acid cycle, and beta oxidation, produce the reduced coenzyme NADH. This coenzyme contains electrons that have a high transfer potential; in other words, they will release a large amount of energy upon oxidation. However, the cell does not release this energy all at once, as this would be an uncontrollable reaction. Instead, the electrons are removed from NADH and passed to oxygen through a series of enzymes that each release a small amount of the energy. This set of enzymes, consisting of complexes I through IV, is called the electron transport chain and is found in the inner membrane of the mitochondrion. Succinate is also oxidized by the electron transport chain, but feeds into the pathway at a different point.
In eukaryotes, the enzymes in this electron transport system use the energy released from O2 by NADH to pump protons across the inner membrane of the mitochondrion. This causes protons to build up in the intermembrane space, and generates an electrochemical gradient across the membrane. The energy stored in this potential is then used by ATP synthase to produce ATP. Oxidative phosphorylation in the eukaryotic mitochondrion is the best-understood example of this process. The mitochondrion is present in almost all eukaryotes, with the exception of anaerobic protozoa such as Trichomonas vaginalis that instead reduce protons to hydrogen in a remnant mitochondrion called a hydrogenosome.
| Respiratory enzyme | Redox pair | Midpoint potential |
|---|---|---|
| NADH dehydrogenase | NAD+ / NADH | −0.32 |
| Succinate dehydrogenase | FMN or FAD / FMNH2 or FADH2 | −0.20 |
| Cytochrome bc1 complex | Coenzyme Q10ox / Coenzyme Q10red | +0.06 |
| Cytochrome bc1 complex | Cytochrome box / Cytochrome bred | +0.12 |
| Complex IV | Cytochrome cox / Cytochrome cred | +0.22 |
| Complex IV | Cytochrome aox / Cytochrome ared | +0.29 |
| Complex IV | O2 / HO− | +0.82 |
| Conditions: pH = 7 |
NADH-coenzyme Q oxidoreductase (complex I)
NADH-coenzyme Q oxidoreductase, also known as NADH dehydrogenase or complex I, is the first protein in the electron transport chain. Complex I is a giant enzyme with the mammalian complex I having 46 subunits and a molecular mass of about . The structure is known in detail only from a bacterium; in most organisms the complex resembles a boot with a large "ball" poking out from the membrane into the mitochondrion. The genes that encode the individual proteins are contained in both the cell nucleus and the mitochondrial genome, as is the case for many enzymes present in the mitochondrion.
The reaction that is catalyzed by this enzyme is the two electron oxidation of NADH by coenzyme Q10 or ubiquinone (represented as Q in the equation below), a lipid-soluble quinone that is found in the mitochondrion membrane:
The start of the reaction, and indeed of the entire electron chain, is the binding of a NADH molecule to complex I and the donation of two electrons. The electrons enter complex I via a prosthetic group attached to the complex, flavin mononucleotide (FMN). The addition of electrons to FMN converts it to its reduced form, FMNH2. The electrons are then transferred through a series of iron–sulfur clusters: the second kind of prosthetic group present in the complex. There are both [2Fe–2S] and [4Fe–4S] iron–sulfur clusters in complex I.
As the electrons pass through this complex, four protons are pumped from the matrix into the intermembrane space. Exactly how this occurs is unclear, but it seems to involve conformational changes in complex I that cause the protein to bind protons on the N-side of the membrane and release them on the P-side of the membrane. Finally, the electrons are transferred from the chain of iron–sulfur clusters to a ubiquinone molecule in the membrane. Reduction of ubiquinone also contributes to the generation of a proton gradient, as two protons are taken up from the matrix as it is reduced to ubiquinol (QH2).
Succinate-Q oxidoreductase (complex II)

Succinate-Q oxidoreductase, also known as complex II or succinate dehydrogenase, is a second entry point to the electron transport chain. It is unusual because it is the only enzyme that is part of both the citric acid cycle and the electron transport chain. Complex II consists of four protein subunits and contains a bound flavin adenine dinucleotide (FAD) cofactor, iron–sulfur clusters, and a heme group that does not participate in electron transfer to coenzyme Q, but is believed to be important in decreasing production of reactive oxygen species. It oxidizes succinate to fumarate and reduces ubiquinone. As this reaction releases less energy than the oxidation of NADH, complex II does not transport protons across the membrane and does not contribute to the proton gradient.
In some eukaryotes, such as the parasitic worm Ascaris suum, an enzyme similar to complex II, fumarate reductase (menaquinol:fumarate oxidoreductase, or QFR), operates in reverse to oxidize ubiquinol and reduce fumarate. This allows the worm to survive in the anaerobic environment of the large intestine, carrying out anaerobic oxidative phosphorylation with fumarate as the electron acceptor. Another unconventional function of complex II is seen in the malaria parasite Plasmodium falciparum. Here, the reversed action of complex II as an oxidase is important in regenerating ubiquinol, which the parasite uses in an unusual form of pyrimidine biosynthesis.
Electron transfer flavoprotein-Q oxidoreductase
Electron transfer flavoprotein-ubiquinone oxidoreductase (ETF-Q oxidoreductase), also known as electron transferring-flavoprotein dehydrogenase, is a third entry point to the electron transport chain. It is an enzyme that accepts electrons from electron-transferring flavoprotein in the mitochondrial matrix, and uses these electrons to reduce ubiquinone. This enzyme contains a flavin and a [4Fe–4S] cluster, but, unlike the other respiratory complexes, it attaches to the surface of the membrane and does not cross the lipid bilayer.
In mammals, this metabolic pathway is important in beta oxidation of fatty acids and catabolism of amino acids and choline, as it accepts electrons from multiple acetyl-CoA dehydrogenases. In plants, ETF-Q oxidoreductase is also important in the metabolic responses that allow survival in extended periods of darkness.
Q-cytochrome c oxidoreductase (complex III)

Q-cytochrome c oxidoreductase is also known as cytochrome c reductase, cytochrome bc1 complex, or simply complex III. In mammals, this enzyme is a dimer, with each subunit complex containing 11 protein subunits, an [2Fe-2S] iron–sulfur cluster and three cytochromes: one cytochrome c1 and two b cytochromes. A cytochrome is a kind of electron-transferring protein that contains at least one heme group. The iron atoms inside complex III's heme groups alternate between a reduced ferrous (+2) and oxidized ferric (+3) state as the electrons are transferred through the protein.
The reaction catalyzed by complex III is the oxidation of one molecule of ubiquinol and the reduction of two molecules of cytochrome c, a heme protein loosely associated with the mitochondrion. Unlike coenzyme Q, which carries two electrons, cytochrome c carries only one electron.
As only one of the electrons can be transferred from the QH2 donor to a cytochrome c acceptor at a time, the reaction mechanism of complex III is more elaborate than those of the other respiratory complexes, and occurs in two steps called the Q cycle. In the first step, the enzyme binds three substrates, first, QH2, which is then oxidized, with one electron being passed to the second substrate, cytochrome c. The two protons released from QH2 pass into the intermembrane space. The third substrate is Q, which accepts the second electron from the QH2 and is reduced to Q.−, which is the ubisemiquinone free radical. The first two substrates are released, but this ubisemiquinone intermediate remains bound. In the second step, a second molecule of QH2 is bound and again passes its first electron to a cytochrome c acceptor. The second electron is passed to the bound ubisemiquinone, reducing it to QH2 as it gains two protons from the mitochondrial matrix. This QH2 is then released from the enzyme.
As coenzyme Q is reduced to ubiquinol on the inner side of the membrane and oxidized to ubiquinone on the other, a net transfer of protons across the membrane occurs, adding to the proton gradient. The rather complex two-step mechanism by which this occurs is important, as it increases the efficiency of proton transfer. If, instead of the Q cycle, one molecule of QH2 were used to directly reduce two molecules of cytochrome c, the efficiency would be halved, with only one proton transferred per cytochrome c reduced.
Cytochrome c oxidase (complex IV)

Cytochrome c oxidase, also known as complex IV, is the final protein complex in the electron transport chain. The mammalian enzyme has an extremely complicated structure and contains 13 subunits, two heme groups, as well as multiple metal ion cofactors – in all, three atoms of copper, one of magnesium and one of zinc.
This enzyme mediates the final reaction in the electron transport chain and transfers electrons to oxygen and hydrogen (protons), while pumping protons across the membrane. The final electron acceptor oxygen is reduced to water in this step. Both the direct pumping of protons and the consumption of matrix protons in the reduction of oxygen contribute to the proton gradient. The reaction catalyzed is the oxidation of cytochrome c and the reduction of oxygen:
Alternative reductases and oxidases
Many eukaryotic organisms have electron transport chains that differ from the much-studied mammalian enzymes described above. For example, plants have alternative NADH oxidases, which oxidize NADH in the cytosol rather than in the mitochondrial matrix, and pass these electrons to the ubiquinone pool. These enzymes do not transport protons, and, therefore, reduce ubiquinone without altering the electrochemical gradient across the inner membrane.
Another example of a divergent electron transport chain is the alternative oxidase, which is found in plants, as well as some fungi, protists, and possibly some animals. This enzyme transfers electrons directly from ubiquinol to oxygen.
The electron transport pathways produced by these alternative NADH and ubiquinone oxidases have lower ATP yields than the full pathway. The advantages produced by a shortened pathway are not entirely clear. However, the alternative oxidase is produced in response to stresses such as cold, reactive oxygen species, and infection by pathogens, as well as other factors that inhibit the full electron transport chain. Alternative pathways might, therefore, enhance an organism's resistance to injury, by reducing oxidative stress.
Organization of complexes
The original model for how the respiratory chain complexes are organized was that they diffuse freely and independently in the mitochondrial membrane. However, recent data suggest that the complexes might form higher-order structures called supercomplexes or "respirasomes". In this model, the various complexes exist as organized sets of interacting enzymes. These associations might allow channeling of substrates between the various enzyme complexes, increasing the rate and efficiency of electron transfer. Within such mammalian supercomplexes, some components would be present in higher amounts than others, with some data suggesting a ratio between complexes I/II/III/IV and the ATP synthase of approximately 1:1:3:7:4. However, the debate over this supercomplex hypothesis is not completely resolved, as some data do not appear to fit with this model.
Prokaryotic electron transport chains
In contrast to the general similarity in structure and function of the electron transport chains in eukaryotes, bacteria and archaea possess a large variety of electron-transfer enzymes. These use an equally wide set of chemicals as substrates. In common with eukaryotes, prokaryotic electron transport uses the energy released from the oxidation of a substrate to pump ions across a membrane and generate an electrochemical gradient. For bacteria, oxidative phosphorylation is understood in most detail in Escherichia coli, while archaeal systems are, at present, poorly understood.
The main difference between eukaryotic and prokaryotic oxidative phosphorylation is that bacteria and archaea use many different substances to donate or accept electrons. This allows prokaryotes to grow under a wide variety of environmental conditions. In E. coli, for example, oxidative phosphorylation can be driven by a large number of pairs of reducing agents and oxidizing agents, which are listed below. The midpoint potential of a chemical measures how much energy is released when it is oxidized or reduced, with reducing agents having negative potentials and oxidizing agents positive potentials.
| Respiratory enzyme | Redox pair | Midpoint potential |
|---|---|---|
| Ubiquinol oxidase | Oxygen / Water | +0.82 |
| Nitrate reductase | Nitrate / Nitrite | +0.42 |
| Nitrite reductase | Nitrite / Ammonia | +0.36 |
| Dimethyl sulfoxide reductase | DMSO / DMS | +0.16 |
| Trimethylamine N-oxide reductase | TMAO / TMA | +0.13 |
| Fumarate reductase | Fumarate / Succinate | +0.03 |
| Succinate dehydrogenase | Fumarate / Succinate | +0.03 |
| Glucose dehydrogenase | Gluconate / Glucose | −0.14 |
| Glycerol-3-phosphate dehydrogenase | DHAP / Gly-3-P | −0.19 |
| Lactate dehydrogenase | Pyruvate / Lactate | −0.19 |
| NADH dehydrogenase | NAD+ / NADH | −0.32 |
| Hydrogenase | Proton / Hydrogen | −0.42 |
| Formate dehydrogenase | Bicarbonate / Formate | −0.43 |
| D-amino acid dehydrogenase | 2-oxoacid + ammonia / D-amino acid | ? |
| Pyruvate oxidase | Acetate + Carbon dioxide / Pyruvate | ? |
As shown above, E. coli can grow with reducing agents such as formate, hydrogen, or lactate as electron donors, and nitrate, DMSO, or oxygen as acceptors.
Some prokaryotes use redox pairs that have only a small difference in midpoint potential. For example, nitrifying bacteria such as Nitrobacter oxidize nitrite to nitrate, donating the electrons to oxygen. The small amount of energy released in this reaction is enough to pump protons and generate ATP, but not enough to produce NADH or NADPH directly for use in anabolism. This problem is solved by using a nitrite oxidoreductase to produce enough proton-motive force to run part of the electron transport chain in reverse, causing complex I to generate NADH.
Prokaryotes control their use of these electron donors and acceptors by varying which enzymes are produced in response to environmental conditions. This flexibility is possible because different oxidases and reductases use the same ubiquinone pool. This allows many combinations of enzymes to function together, linked by the common ubiquinol intermediate. These respiratory chains therefore have a modular design, with easily interchangeable sets of enzyme systems.
In addition to this metabolic diversity, prokaryotes also possess a range of isozymes – different enzymes that catalyze the same reaction. For example, in E. coli, there are two different types of ubiquinol oxidase using oxygen as an electron acceptor. Under highly aerobic conditions, the cell uses an oxidase with a low affinity for oxygen that can transport two protons per electron. However, if levels of oxygen fall, they switch to an oxidase that transfers only one proton per electron, but has a high affinity for oxygen.
ATP synthase (complex V)
ATP synthase, also called complex V, is the final enzyme in the oxidative phosphorylation pathway. This enzyme is found in all forms of life and functions in the same way in both prokaryotes and eukaryotes. The enzyme uses the energy stored in a proton gradient across a membrane to drive the synthesis of ATP from ADP and phosphate (Pi). Estimates of the number of protons required to synthesize one ATP have ranged from three to four, with some suggesting cells can vary this ratio, to suit different conditions.
This phosphorylation reaction is an equilibrium, which can be shifted by altering the proton-motive force. In the absence of a proton-motive force, the ATP synthase reaction will run from right to left, hydrolyzing ATP and pumping protons out of the matrix across the membrane. However, when the proton-motive force is high, the reaction is forced to run in the opposite direction; it proceeds from left to right, allowing protons to flow down their concentration gradient and turning ADP into ATP.
ATP synthase is a massive protein complex with a mushroom-like shape. The mammalian enzyme complex contains 16 subunits and has a mass of approximately 600 kilodaltons. The portion embedded within the membrane is called FO and contains a ring of c subunits and the proton channel. The stalk and the ball-shaped headpiece is called F1 and is the site of ATP synthesis. The ball-shaped complex at the end of the F1 portion contains six proteins of two different kinds (three α subunits and three β subunits), whereas the "stalk" consists of one protein: the γ subunit, with the tip of the stalk extending into the ball of α and β subunits. Both the α and β subunits bind nucleotides, but only the β subunits catalyze the ATP synthesis reaction. Reaching along the side of the F1 portion and back into the membrane is a long rod-like subunit that anchors the α and β subunits into the base of the enzyme.
As protons cross the membrane through the channel in the base of ATP synthase, the FO proton-driven motor rotates. Rotation might be caused by changes in the ionization of amino acids in the ring of c subunits causing electrostatic interactions that propel the ring of c subunits past the proton channel. This rotating ring in turn drives the rotation of the central axle (the γ subunit stalk) within the α and β subunits. The α and β subunits are prevented from rotating themselves by the side-arm, which acts as a stator. This movement of the tip of the γ subunit within the ball of α and β subunits provides the energy for the active sites in the β subunits to undergo a cycle of movements that produces and then releases ATP.
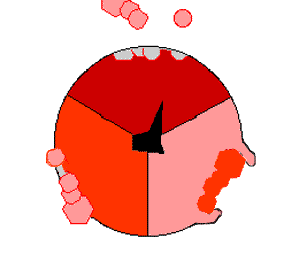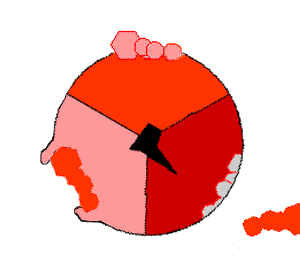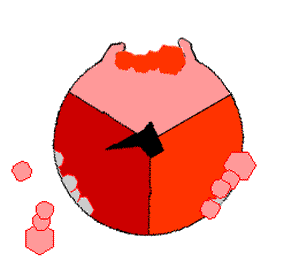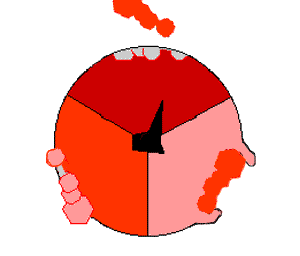
This ATP synthesis reaction is called the binding change mechanism and involves the active site of a β subunit cycling between three states. In the "open" state, ADP and phosphate enter the active site (shown in brown in the diagram). The protein then closes up around the molecules and binds them loosely – the "loose" state (shown in red). The enzyme then changes shape again and forces these molecules together, with the active site in the resulting "tight" state (shown in pink) binding the newly produced ATP molecule with very high affinity. Finally, the active site cycles back to the open state, releasing ATP and binding more ADP and phosphate, ready for the next cycle.
In some bacteria and archaea, ATP synthesis is driven by the movement of sodium ions through the cell membrane, rather than the movement of protons. Archaea such as Methanococcus also contain the A1Ao synthase, a form of the enzyme that contains additional proteins with little similarity in sequence to other bacterial and eukaryotic ATP synthase subunits. It is possible that, in some species, the A1Ao form of the enzyme is a specialized sodium-driven ATP synthase, but this might not be true in all cases.
Reactive oxygen species
Molecular oxygen is a good terminal electron acceptor because it is a strong oxidizing agent. The reduction of oxygen does involve potentially harmful intermediates. Although the transfer of four electrons and four protons reduces oxygen to water, which is harmless, transfer of one or two electrons produces superoxide or peroxide anions, which are dangerously reactive.
} -[\ce{e^-}] \underset{Peroxide}{O2^{2-}}|}}
These reactive oxygen species and their reaction products, such as the hydroxyl radical, are very harmful to cells, as they oxidize proteins and cause mutations in DNA. This cellular damage may contribute to disease and is proposed as one cause of aging.
The cytochrome c oxidase complex is highly efficient at reducing oxygen to water, and it releases very few partly reduced intermediates; however small amounts of superoxide anion and peroxide are produced by the electron transport chain. Particularly important is the reduction of coenzyme Q in complex III, as a highly reactive ubisemiquinone free radical is formed as an intermediate in the Q cycle. This unstable species can lead to electron "leakage" when electrons transfer directly to oxygen, forming superoxide. As the production of reactive oxygen species by these proton-pumping complexes is greatest at high membrane potentials, it has been proposed that mitochondria regulate their activity to maintain the membrane potential within a narrow range that balances ATP production against oxidant generation. For instance, oxidants can activate uncoupling proteins that reduce membrane potential.
To counteract these reactive oxygen species, cells contain numerous antioxidant systems, including antioxidant vitamins such as vitamin C and vitamin E, and antioxidant enzymes such as superoxide dismutase, catalase, and peroxidases, which detoxify the reactive species, limiting damage to the cell.
In hypoxic/anoxic conditions
As oxygen is fundamental for oxidative phosphorylation, a shortage in O2 level can alter ATP production rates. The proton motive force and ATP production can be maintained by intracellular acidosis. Cytosolic protons that have accumulated with ATP hydrolysis and lactic acidosis can freely diffuse across the mitochondrial outer-membrane and acidify the inter-membrane space, hence directly contributing to the proton motive force and ATP production.
When exposed to hypoxia/anoxia (no oxygen), most animals will see damage done to their mitochondria. From some species, these conditions can happen due to environmental variables, such as low tides, low temperatures, or general living conditions, like living in a hypoxic underground burrow. In humans, these conditions are commonly met in medical emergencies such as strokes, ischemia, and asphyxia.
Despite this, or perhaps due to it, some species have developed their own defense mechanisms against anoxia/hypoxia, as well as during reperfusion/reoxygenation. These mechanisms are diverse and differ between endotherms and ectotherms and can differ even at the species level.
Endotherms
Hypoxia/anoxia intolerance
Most mammals and birds are intolerant to low/no oxygen conditions. For the heart, in the absence of oxygen, the first four complexes of the electron transport chain decrease in activity. This will lead to protons leaking through the inner mitochondrial membrane without complexes I, III, and IV pushing protons back through to maintain the proton gradient. There is also electron leak (an event where electrons leak out of the electron transport chain), which happens because NADH dehydrogenase within Complex I becomes damaged, which allows for the production of ROS (reactive oxygen species) during ischemia. This will lead to the reversing of Complex V, which forces protons from the matrix back into the inner membrane space, against their concentration gradient. Forcing protons against their concentration gradient requires energy, so Complex V uses up ATP as an energy source.
Reoxygenation of intolerant animals
When oxygen re-enters the system, animals are faced with a different set of problems. Since ATP was used up during the anoxic period, it leads to a lack of ADP within the system. This is due to ADP's natural degradation into AMP, resulting in ADP being drained from the system. With no ADP in the system, Complex V is unable to start, meaning the protons will not flow through it to enter the matrix. Due to Complex V's reversal during anoxia, the proton gradient has become hyperpolarized (where the proton gradient is highly positively charged). Another factor in this problem is that succinate built up during anoxia, so when oxygen is reintroduced, succinate donates electrons to Complex II. The hyperpolarized gradient and succinate buildup leads to reverse electron transport, causing oxidative stress, which can lead to cellular damage and diseases.
Hypoxia/anoxia tolerance
The naked mole rat (Heterocephalus glaber) is a hypoxia-tolerant species that sleeps in deep burrows and in large colonies. The depth of these burrows reduces access to oxygen, and sleeping in large groups will deplete the area of oxygen quicker than usual, leading to hypoxia. The naked mole rat has the unique ability to survive low oxygen conditions for no less than several hours, and zero oxygen conditions for 18 minutes. One of the ways of combatting hypoxia in the brain is decreasing the reliance on oxygen for ATP production, achieved by decreased respiration rates and proton leak.
Reoxygenation of tolerant animals
Hypoxia/anoxia tolerant species handle ROS production during reoxygenation better than the intolerant. In the cortex of the naked mole rats, they show better homeostasis of ROS production than intolerant species and seem to lack the burst of ROS that typically comes with reoxygenation.
Ectotherms
Hypoxia/anoxia intolerance
Research on intolerant ectotherms is more limited than on tolerant ectotherms and intolerant endotherms, but it is shown that anoxia/hypoxia intolerance is different in terms for how long the intolerant survive as opposed to the tolerant between endotherms and ectotherms. While intolerant endotherms only last minutes, intolerant ectotherms can last hours, such as subtidal scallops (Argopecten irradians). This difference in intolerance could be due to a couple of different factors. One advantage is that the ectothermic inner mitochondrial membrane is less leaky, so less protons will leak through the inner membrane due to differences in the phospholipid bilayer composition. Another advantage ectotherms tend to have in this category is an ability for their mitochondria to properly function in a wide range of temperatures, such as the western fence lizard (Sceloporus occidentalis). While western fence lizards are not considered a hypoxia-tolerant animal, they still showed less temperature sensitivity in their mitochondria than mice mitochondria.
Reoxygenation of intolerant animals
While it is unclear how reoxygenation affects intolerant ectotherms at the mitochondrial level, there is some research showing how some of them respond. In the hypoxia-sensitive shovelnose ray (Aptychotrema rostrata), it is shown that ROS production is lower upon reoxygenation compared to rays only exposed to normoxia (normal oxygen levels). This differs from the hypoxia-sensitive endotherm, which would see an increase in ROS production. However, the ray's levels were still higher than the more hypoxia-tolerant Epaulette shark (Hemiscyllum ocellatum), which potentially sees hypoxia due to the bouts of low tides that can be seen in reef platforms. Subtidal scallops will see both a decrease in maximal respiration and a depolarization of the membrane during reoxygenation.
Hypoxia/anoxia tolerance
Hypoxia/Anoxia tolerant ectotherms have shown unique strategies for surviving anoxia. Pond turtles, such as the painted turtle (Chrysemys picta bellii), will experience anoxia during winter while they overwinter at the bottom of frozen ponds. In their cardiac mitochondria, the reversing of Complex V, the usage of ATP, and the build-up of succinate are all prevented during anoxia. Crucian carps (Carassius carassius) also overwinter in frozen ponds and show no loss membrane potential in their cardiac mitochondria during anoxia, but this relies on complexes I and III to be active.
Reoxygenation of tolerant animals
Pond turtles are able to completely avoid ROS production upon reoxygenation. However, crucian carp cannot and are unable to prevent the death of brain cells upon reoxygenation.
Inhibitors
There are several well-known drugs and toxins that inhibit oxidative phosphorylation. Although any one of these toxins inhibits only one enzyme in the electron transport chain, inhibition of any step in this process will halt the rest of the process. For example, if oligomycin inhibits ATP synthase, protons cannot pass back into the mitochondrion. As a result, the proton pumps are unable to operate, as the gradient becomes too strong for them to overcome. NADH is then no longer oxidized and the citric acid cycle ceases to operate because the concentration of NAD+ falls below the concentration that these enzymes can use.
Many site-specific inhibitors of the electron transport chain have contributed to the present knowledge of mitochondrial respiration. Synthesis of ATP is also dependent on the electron transport chain, so all site-specific inhibitors also inhibit ATP formation. The fish poison rotenone, the barbiturate drug amytal, and the antibiotic piericidin A inhibit NADH and coenzyme Q.
Carbon monoxide, cyanide, hydrogen sulfide and azide effectively inhibit cytochrome oxidase. Carbon monoxide reacts with the reduced form of the cytochrome while cyanide and azide react with the oxidised form. An antibiotic, antimycin A, and British anti-Lewisite, an antidote used against chemical weapons, are the two important inhibitors of the site between cytochrome B and C1.
| Compounds | Use | Site of action | Effect on oxidative phosphorylation | |
|---|---|---|---|---|
| Cyanide | ||||
| Carbon monoxide | ||||
| Azide | ||||
| Hydrogen sulfide | Poisons | Complex IV | Inhibit the electron transport chain by binding more strongly than oxygen to the Fe–Cu center in cytochrome c oxidase, preventing the reduction of oxygen. | |
| Oligomycin | Antibiotic | Complex V | Inhibits ATP synthase by blocking the flow of protons through the Fo subunit. | |
| CCCP | ||||
| 2,4-Dinitrophenol | Poisons, weight-lossDNP was extensively used as an anti-obesity medication in the 1930s but was ultimately discontinued due to its dangerous side effects. However, illicit use of the drug for this purpose continues today. See 2,4-Dinitrophenol#Dieting aid for more information. | Inner membrane | Ionophores that disrupt the proton gradient by carrying protons across a membrane. This ionophore uncouples proton pumping from ATP synthesis because it carries protons across the inner mitochondrial membrane. | |
| Rotenone | Pesticide | Complex I | Prevents the transfer of electrons from complex I to ubiquinone by blocking the ubiquinone-binding site. | |
| Malonate and oxaloacetate | Poisons | Complex II | Competitive inhibitors of succinate dehydrogenase (complex II). | |
| Antimycin A | Piscicide | Complex III | Binds to the Qi site of cytochrome c reductase, thereby inhibiting the oxidation of ubiquinol. |
Not all inhibitors of oxidative phosphorylation are toxins. In brown adipose tissue, regulated proton channels called uncoupling proteins can uncouple respiration from ATP synthesis. This rapid respiration produces heat, and is particularly important as a way of maintaining body temperature for hibernating animals, although these proteins may also have a more general function in cells' responses to stress.
History
The field of oxidative phosphorylation began with the report in 1906 by Arthur Harden of a vital role for phosphate in cellular fermentation, but initially only sugar phosphates were known to be involved. However, in the early 1940s, the link between the oxidation of sugars and the generation of ATP was firmly established by Herman Kalckar, confirming the central role of ATP in energy transfer that had been proposed by Fritz Albert Lipmann in 1941. Later, in 1949, Morris Friedkin and Albert L. Lehninger proved that the coenzyme NADH linked metabolic pathways such as the citric acid cycle and the synthesis of ATP. The term oxidative phosphorylation was coined by in 1939.
For another twenty years, the mechanism by which ATP is generated remained mysterious, with scientists searching for an elusive "high-energy intermediate" that would link oxidation and phosphorylation reactions. This puzzle was solved by Peter D. Mitchell with the publication of the chemiosmotic theory in 1961. At first, this proposal was highly controversial, but it was slowly accepted and Mitchell was awarded a Nobel prize in 1978. Subsequent research concentrated on purifying and characterizing the enzymes involved, with major contributions being made by David E. Green on the complexes of the electron-transport chain, as well as Efraim Racker on the ATP synthase. A critical step towards solving the mechanism of the ATP synthase was provided by Paul D. Boyer, by his development in 1973 of the "binding change" mechanism, followed by his radical proposal of rotational catalysis in 1982. More recent work has included structural studies on the enzymes involved in oxidative phosphorylation by John E. Walker, with Walker and Boyer being awarded a Nobel Prize in 1997.
Notes
References
References
- Voet, Donald. (2004). "Biochemistry". Wiley.
- (January 1967). "Chemiosmotic hypothesis of oxidative phosphorylation". Nature.
- (January 2000). "Crucial role of the membrane potential for ATP synthesis by F(1)F(o) ATP synthases". The Journal of Experimental Biology.
- (2001). "Structures and proton-pumping strategies of mitochondrial respiratory enzymes". Annual Review of Biophysics and Biomolecular Structure.
- (December 2003). "The molecular machinery of Keilin's respiratory chain". Biochemical Society Transactions.
- (September 1995). "Mitochondrial proton conductance and H+/O ratio are independent of electron transport rate in isolated hepatocytes". The Biochemical Journal.
- (1985). "The structure, function and evolution of cytochromes". Progress in Biophysics and Molecular Biology.
- (December 1983). "Why do c-type cytochromes exist?". FEBS Letters.
- (December 2001). "Biochemical functions of coenzyme Q10". Journal of the American College of Nutrition.
- (December 1979). "Keilin's respiratory chain concept and its chemiosmotic consequences". Science.
- (August 1999). "Microbial ubiquinones: multiple roles in respiration, gene regulation and oxidative stress management". Microbiology.
- (2005). "Structure, function, and formation of biological iron-sulfur clusters". Annual Review of Biochemistry.
- (November 1999). "Natural engineering principles of electron tunnelling in biological oxidation-reduction". Nature.
- (December 2004). "Electrical circuitry in biology: emerging principles from protein structure". Current Opinion in Structural Biology.
- (March 2005). "An anaerobic mitochondrion that produces hydrogen". Nature.
- Medical CHEMISTRY Compendium. By Anders Overgaard Pedersen and Henning Nielsen. Aarhus University. 2008
- (June 2005). "Energy transduction by respiratory complex I--an evaluation of current knowledge". Biochemical Society Transactions.
- (2006). "Mitochondrial Complex I: structural and functional aspects". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (March 2006). "Structure of the hydrophilic domain of respiratory complex I from Thermus thermophilus". Science.
- (May 2010). "The architecture of respiratory complex I". Nature.
- (February 2007). "Projection structure of the membrane domain of Escherichia coli respiratory complex I at 8 A resolution". Journal of Molecular Biology.
- (January 2004). "The gross structure of the respiratory complex I: a Lego System". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (December 2009). "Towards the molecular mechanism of respiratory complex I". The Biochemical Journal.
- (2003). "Function and structure of complex II of the respiratory chain". Annual Review of Biochemistry.
- (January 2003). "Architecture of succinate dehydrogenase and reactive oxygen species generation". Science.
- (April 2004). "Complex II from a structural perspective". Current Protein & Peptide Science.
- (January 2002). "Role of complex II in anaerobic respiration of the parasite mitochondria from Ascaris suum and Plasmodium falciparum". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (March 2007). "Specific role of mitochondrial electron transport in blood-stage Plasmodium falciparum". Nature.
- (February 1987). "Reactions of electron-transfer flavoprotein and electron-transfer flavoprotein: ubiquinone oxidoreductase". The Biochemical Journal.
- (October 2006). "Structure of electron transfer flavoprotein-ubiquinone oxidoreductase and electron transfer to the mitochondrial ubiquinone pool". Proceedings of the National Academy of Sciences of the United States of America.
- (January 1983). "Separation and properties of five distinct acyl-CoA dehydrogenases from rat liver mitochondria. Identification of a new 2-methyl branched chain acyl-CoA dehydrogenase". The Journal of Biological Chemistry.
- (December 1977). "A new iron-sulfur flavoprotein of the respiratory chain. A component of the fatty acid beta oxidation pathway". The Journal of Biological Chemistry.
- (September 2005). "The critical role of Arabidopsis electron-transfer flavoprotein:ubiquinone oxidoreductase during dark-induced starvation". The Plant Cell.
- (2000). "Structure and function of cytochrome bc complexes". Annual Review of Biochemistry.
- (2004). "The cytochrome bc1 complex: function in the context of structure". Annual Review of Physiology.
- (July 1998). "Complete structure of the 11-subunit bovine mitochondrial cytochrome bc1 complex". Science.
- (July 1990). "The protonmotive Q cycle. Energy transduction by coupling of proton translocation to electron transfer by the cytochrome bc1 complex". The Journal of Biological Chemistry.
- (June 2003). "Protonmotive pathways and mechanisms in the cytochrome bc1 complex". FEBS Letters.
- (August 1994). "The cytochrome oxidase superfamily of redox-driven proton pumps". Trends in Biochemical Sciences.
- (May 1996). "The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A". Science.
- (2006). "Proton pumping mechanism of bovine heart cytochrome c oxidase". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (2004). "Alternative NAD(P)H dehydrogenases of plant mitochondria". Annual Review of Plant Biology.
- (September 1996). "Purification and characterization of a 43-kDa rotenone-insensitive NADH dehydrogenase from plant mitochondria". The Journal of Biological Chemistry.
- (June 2004). "Branched mitochondrial electron transport in the Animalia: presence of alternative oxidase in several animal phyla". IUBMB Life.
- (June 1998). "Alternative oxidase in the branched mitochondrial respiratory network: an overview on structure, function, regulation, and role". Brazilian Journal of Medical and Biological Research = Revista Brasileira de Pesquisas Medicas e Biologicas.
- (August 1991). "The regulation and nature of the cyanide-resistant alternative oxidase of plant mitochondria". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (June 1997). "ALTERNATIVE OXIDASE: From Gene to Function". Annual Review of Plant Physiology and Plant Molecular Biology.
- (December 1997). "Transcript levels of tandem-arranged alternative oxidase genes in rice are increased by low temperature". Gene.
- (July 1999). "The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells". Proceedings of the National Academy of Sciences of the United States of America.
- (December 2001). "A critical appraisal of the mitochondrial coenzyme Q pool". FEBS Letters.
- (April 2007). "A structural model of the cytochrome C reductase/oxidase supercomplex from yeast mitochondria". The Journal of Biological Chemistry.
- (April 2000). "Supercomplexes in the respiratory chains of yeast and mammalian mitochondria". The EMBO Journal.
- (September 2002). "Respiratory chain supercomplexes of mitochondria and bacteria". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (October 2001). "The ratio of oxidative phosphorylation complexes I-V in bovine heart mitochondria and the composition of respiratory chain supercomplexes". The Journal of Biological Chemistry.
- (May 1984). "Relationship between lateral diffusion, collision frequency, and electron transfer of mitochondrial inner membrane oxidation-reduction components". Proceedings of the National Academy of Sciences of the United States of America.
- (January 1999). "Post-Viking microbiology: new approaches, new data, new insights". Origins of Life and Evolution of the Biosphere.
- (September 1999). "Bioenergetics of the Archaea". Microbiology and Molecular Biology Reviews.
- (September 1984). "The respiratory chains of Escherichia coli". Microbiological Reviews.
- (July 1997). "Alternative respiratory pathways of Escherichia coli: energetics and transcriptional regulation in response to electron acceptors". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (January 2002). "Succinate dehydrogenase and fumarate reductase from Escherichia coli". Biochimica et Biophysica Acta (BBA) - Bioenergetics.
- (1990). "Energy conservation in Nitrobacter". FEMS Microbiology Letters.
- (March 2006). "Genome sequence of the chemolithoautotrophic nitrite-oxidizing bacterium Nitrobacter winogradskyi Nb-255". Applied and Environmental Microbiology.
- (December 1988). "The nitrite oxidizing system of Nitrobacter winogradskyi". FEMS Microbiology Reviews.
- (July 1993). "Adaptation of Escherichia coli to redox environments by gene expression". Molecular Microbiology.
- (May 1993). "Energetic efficiency of Escherichia coli: effects of mutations in components of the aerobic respiratory chain". Journal of Bacteriology.
- (1997). "The ATP synthase--a splendid molecular machine". Annual Review of Biochemistry.
- (February 1996). "The H+/ATP coupling ratio of the ATP synthase from thiol-modulated chloroplasts and two cyanobacterial strains is four". FEBS Letters.
- (September 2001). "ATP synthase--a marvellous rotary engine of the cell". Nature Reviews. Molecular Cell Biology.
- (June 1998). "Effects of carbon source on expression of F0 genes and on the stoichiometry of the c subunit in the F1F0 ATPase of Escherichia coli". Journal of Bacteriology.
- (January 2000). "The cellular biology of proton-motive force generation by V-ATPases". The Journal of Experimental Biology.
- (December 2003). "Structure of the mitochondrial ATP synthase by electron cryomicroscopy". The EMBO Journal.
- (April 2000). "Structural model of F1-ATPase and the implications for rotary catalysis". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences.
- (January 2001). "The rotary machine in the cell, ATP synthase". The Journal of Biological Chemistry.
- (March 2002). "Mechanism of the F(1)F(0)-type ATP synthase, a biological rotary motor". Trends in Biochemical Sciences.
- (March 2006). "Catalytic and mechanical cycles in F-ATP synthases. Fourth in the Cycles Review Series". EMBO Reports.
- (October 1982). "Catalytic site cooperativity of beef heart mitochondrial F1 adenosine triphosphatase. Correlations of initial velocity, bound intermediate, and oxygen exchange measurements with an alternating three-site model". The Journal of Biological Chemistry.
- (1994). "Bacterial sodium ion-coupled energetics". Antonie van Leeuwenhoek.
- (May 1994). "Delta mu Na+ drives the synthesis of ATP via an delta mu Na(+)-translocating F1F0-ATP synthase in membrane vesicles of the archaeon Methanosarcina mazei Gö1". Journal of Bacteriology.
- (February 2004). "An exceptional variability in the motor of archael A1A0 ATPases: from multimeric to monomeric rotors comprising 6-13 ion binding sites". Journal of Bioenergetics and Biomembranes.
- (1995). "Oxidative stress: the paradox of aerobic life". Biochemical Society Symposium.
- (December 2006). "Theories of biological aging: genes, proteins, and free radicals". Free Radical Research.
- (2007). "Free radicals and antioxidants in normal physiological functions and human disease". The International Journal of Biochemistry & Cell Biology.
- (October 2000). "Mitochondria, oxygen free radicals, disease and ageing". Trends in Biochemical Sciences.
- (November 2000). "Oxidants, oxidative stress and the biology of ageing". Nature.
- (March 2010). "New extension of the Mitchell Theory for oxidative phosphorylation in mitochondria of living organisms". Biochimica et Biophysica Acta (BBA) - General Subjects.
- (January 2002). "Superoxide activates mitochondrial uncoupling proteins". Nature.
- (January 2019). "Acidosis Maintains the Function of Brain Mitochondria in Hypoxia-Tolerant Triplefin Fish: A Strategy to Survive Acute Hypoxic Exposure?". Frontiers in Physiology.
- (January 2017). "Mitochondrial Dysfunction and Myocardial Ischemia-Reperfusion: Implications for Novel Therapies". Annual Review of Pharmacology and Toxicology.
- (January 2012). "A radical approach to beating hypoxia: depressed free radical release from heart fibres of the hypoxia-tolerant epaulette shark (Hemiscyllum ocellatum)". Journal of Comparative Physiology B.
- (2022). "Review: A history and perspective of mitochondria in the context of anoxia tolerance". Comparative Biochemistry and Physiology. Part B, Biochemistry & Molecular Biology.
- (February 2018). "Naked mole rat brain mitochondria electron transport system flux and H+ leak are reduced during acute hypoxia". The Journal of Experimental Biology.
- (January 2007). "Modulation of electron transport protects cardiac mitochondria and decreases myocardial injury during ischemia and reperfusion". American Journal of Physiology. Cell Physiology.
- (July 2000). "Mitochondria as ATP consumers: cellular treason in anoxia". Proceedings of the National Academy of Sciences of the United States of America.
- (February 2019). "Metabolic adaptations during extreme anoxia in the turtle heart and their implications for ischemia-reperfusion injury". Scientific Reports.
- (March 2024). "Are reactive oxygen species always bad? Lessons from hypoxic ectotherms". The Journal of Experimental Biology.
- (November 2014). "Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS". Nature.
- (January 2009). "How mitochondria produce reactive oxygen species". The Biochemical Journal.
- (March 2013). "Mitochondria and reactive oxygen species: physiology and pathophysiology". International Journal of Molecular Sciences.
- (2023). "Naked Mole-Rat Cortex Maintains Reactive Oxygen Species Homeostasis During ''In Vitro'' Hypoxia or Ischemia and Reperfusion". Current Neuropharmacology.
- (June 2016). "Intermittent hypoxia leads to functional reorganization of mitochondria and affects cellular bioenergetics in marine molluscs". The Journal of Experimental Biology.
- (September 1999). "Oxygen consumption by mitochondria from an endotherm and an ectotherm". Comparative Biochemistry and Physiology. Part B, Biochemistry & Molecular Biology.
- (September 2013). "Beating oxygen: chronic anoxia exposure reduces mitochondrial F1FO-ATPase activity in turtle (Trachemys scripta) heart". The Journal of Experimental Biology.
- (October 2024). "Maintained mitochondrial integrity without oxygen in the anoxia-tolerant crucian carp". The Journal of Experimental Biology.
- (May 2023). "Low production of mitochondrial reactive oxygen species after anoxia and reoxygenation in turtle hearts". The Journal of Experimental Biology.
- (April 2020). "Metabolic adaptations to anoxia and reoxygenation: New lessons from freshwater turtles and crucian carp". Current Opinion in Endocrine and Metabolic Research.
- (2002). "Biochemistry". Books and Allied.
- (January 1993). "Fourier-transform infrared study of cyanide binding to the Fea3-CuB binuclear site of bovine heart cytochrome c oxidase: implication of the redox-linked conformational change at the binuclear site". Biochemistry.
- (August 1991). "ATP synthase complex from bovine heart mitochondria: the oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimide-sensitive ATPase". Biochimica et Biophysica Acta (BBA) - Biomembranes.
- (1979). "Biomembranes Part F: Bioenergetics: Oxidative Phosphorylation".
- (September 2004). "Inhibitors of the quinone-binding site allow rapid superoxide production from mitochondrial NADH:ubiquinone oxidoreductase (complex I)". The Journal of Biological Chemistry.
- (November 1964). "Studies on Succinate Dehydrogenase. I. Spectral Properties of the Purified Enzyme and Formation of Enzyme-Competitive Inhibitor Complexes". Biochimica et Biophysica Acta (BBA) - Specialized Section on Enzymological Subjects.
- (January 2000). "The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP". The Biochemical Journal.
- (2005). "Plant uncoupling mitochondrial protein and alternative oxidase: energy metabolism and stress". Bioscience Reports.
- (1906). "The alcoholic ferment of yeast-juice". Proceedings of the Royal Society.
- (November 1974). "Origins of the concept oxidative phosphorylation". Molecular and Cellular Biochemistry.
- (1941). "Metabolic generation and utilization of phosphate bond energy". Adv Enzymol.
- (April 1949). "Esterification of inorganic phosphate coupled to electron transport between dihydrodiphosphopyridine nucleotide and oxygen". The Journal of Biological Chemistry.
- (1991). "50 years of biological research--from oxidative phosphorylation to energy requiring transport regulation". Annual Review of Biochemistry.
- (1939). "About phosphorilation mechanism coupled with respiration". [[Biokhimiya]].
- (November 1953). "Mechanism of phosphorylation in the respiratory chain". Nature.
- (July 1961). "Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism". Nature.
- "Peter Mitchell and the Vital Force".
- (1978). "David Keilin's Respiratory Chain Concept and Its Chemiosmotic Consequences". Nobel Foundation.
- (November 1960). "Partial resolution of the enzymes catalyzing oxidative phosphorylation. I. Purification and properties of soluble dinitrophenol-stimulated adenosine triphosphatase". The Journal of Biological Chemistry.
- (October 1973). "A new concept for energy coupling in oxidative phosphorylation based on a molecular explanation of the oxygen exchange reactions". Proceedings of the National Academy of Sciences of the United States of America.
- "The Nobel Prize in Chemistry 1997". Nobel Foundation.
This article was imported from Wikipedia and is available under the Creative Commons Attribution-ShareAlike 4.0 License. Content has been adapted to SurfDoc format. Original contributors can be found on the article history page.
Ask Mako anything about Oxidative phosphorylation — get instant answers, deeper analysis, and related topics.
Research with MakoFree with your Surf account
Create a free account to save articles, ask Mako questions, and organize your research.
Sign up freeThis content may have been generated or modified by AI. CloudSurf Software LLC is not responsible for the accuracy, completeness, or reliability of AI-generated content. Always verify important information from primary sources.
Report