From Surf Wiki (app.surf) — the open knowledge base
Haplogroup E-M35
Human Y-chromosome DNA haplogroup
Human Y-chromosome DNA haplogroup
| Field | Value |
|---|---|
| name | E-M35 |
| origin-place | Middle East: "Both phylogeography and microsatellite variance suggest that E-P2 and its derivative, E-M35, probably originated in eastern Africa." |
| origin-date | 34,800 BP |
| TMRCA | 23,900 BP |
| ancestor | E-M215 |
| descendants | E-V68, E-Z827 |
| mutations | M35 |
|origin-place=Middle East: "Both phylogeography and microsatellite variance suggest that E-P2 and its derivative, E-M35, probably originated in eastern Africa." |origin-date=34,800 BP
E-M35, also known as E1b1b1-M35, is a human Y-chromosome DNA haplogroup. E-M35 has two basal branches, E-V68 and E-Z827. E-V68 and E-Z827 are primarily distributed in North Africa and the Horn of Africa, and occur at lower frequencies in the Middle East, Europe, and Southern Africa.
Origins
In June 2015, Trombetta et al. reported a previously unappreciated large difference in the age between haplogroup E-M215 (38.6 kya; 95% CI 31.4–45.9 kya) and its sub-haplogroup E-M35 (25.0 kya; 95% CI 20.0–30.0 kya) and estimated its origin to be in Horn of Africa, where the node separating the E-V38 and E-M215 branches occurs about 47,500 years ago (95% CI: 41.3–56.8 ka). E-M35 was dated by Batini in 2015 to between 15,400 and 20,500 years ago.
All major sub-branches of E-M35 are thought to have originated in the same general area as the parent clade: in North Africa, the Horn of Africa, or nearby areas of the Near East. Some branches of E-M35 are assumed to have left Africa thousands of years ago, whereas others may have arrived from the Near East. For example, associates the spread of the haplogroup with the Neolithic Revolution, believing that the structure and regional pattern of E-M35 subclades potentially give "reagents with which to infer specific episodes of population histories associated with the Neolithic agricultural expansion". also estimate that E-M78 (called E1b1b1a1 in that paper) has been in Europe longer than 10,000 years. Accordingly, human remains excavated in a Spanish funeral cave dating from approximately 7,000 years ago were shown to be in this haplogroup. Two more E-M78 have been found in the Neolithic Sopot and Lengyel cultures too.
Concerning E-M35 in Europe within this scheme, have remarked that E-M215 seems to represent a late-Pleistocene migration from North Africa to Europe over the Sinai Peninsula in Egypt."Y chromosome data show a signal for a separate late-Pleistocene migration from Africa to Europe via Sinai as evidenced through the distribution of haplogroup E3b lineages, which is not manifested in mtDNA haplogroup distributions." While this proposal remains uncontested, it has more recently been proposed by that there is also evidence for additional migration of E-M215 carrying men directly from North Africa to southwestern Europe, via a maritime route (see below.)
Ancient DNA
Loosdrecht et al. (2018) analysed genome-wide data from seven ancient Iberomaurusian individuals from the Grotte des Pigeons near Taforalt in eastern Morocco. The fossils were directly dated to between 15,100 and 13,900 calibrated years before present. The scientists found that five male specimens with sufficient nuclear DNA preservation all belonged to the E1b1b1a1 (M78) subclade, with one skeleton bearing the E1b1b1a1b1 parent lineage to E-V13. Martiniano et al. (2022) later reassigned all the Taforalt samples to haplogroup E-M78 and none to E-L618, the predecessor to E-V13.
According to Lazaridis et al. (2016), Natufian skeletal remains from the ancient Levant predominantly carried the Y-DNA haplogroup E. Of the five Natufian specimens analysed for paternal lineages, one belonged to CT, one to E-M35, one to E-M215, one to E-Z830. Haplogroup E-M35 was also found the dominant marker among fossils from the ensuing Pre-Pottery Neolithic B culture, with the E-M78 and E-Z830 subclades observed in multiple PPNB specimens (~50%). The scientists suggest that the Levantine early farmers may have spread westward into North Africa, bringing along Western Eurasian and Basal Eurasian ancestral components.
Following Fregel et al. (2017), two Neolithic farmers from the North African site of Ifri n Ammar ou Moussa belonged to haplogroup E-L19. The majority of their genomes traced back to Natufian and PPN ancestry , indicating a recent intrusion into North Africa from the Levant. Their presence is likely linked to the spread of pastoral technologies from the Levant into North Africa.
Haplogroup E-M35 has been identified among both Amorite and Akkadian remains, found in the sites of Megiddo, Ebla, and Alalakh. It is also attested among later Semitic populations, such as the Phoenicians who settled throughout the Mediterranean region, where it appears to have been the most prevalent lineage. Additionally, analyses of medieval Andalusian remains indicate that individuals examined for paternal DNA overwhelmingly belonged to the E-M81 subclade.
In Egypt, haplogroup E-V1515 has been identified in the remains of an elite Egyptian individual from the site of Nuerat, dating to the Old Kingdom period, while haplogroup E-V22 has been detected in an ancient Egyptian mummy excavated at the Abusir el-Meleq archaeological site in Middle Egypt, dated to between the late New Kingdom and the Roman era.
Distribution
E-M215 and E-M35 are quite common among Afroasiatic speakers. The linguistic group and carriers of E-M35 lineage have a high probability to have arisen and dispersed together from the Afroasiatic Urheimat. Amongst populations with an Afro-Asiatic speaking history, a significant proportion of Palestinians and Jewish male lineages are E-M35. Haplogroup E-M35, which accounts for approximately 18% to 20% of 18% of Palestinians and Ashkenazi and 8.6% to 30% of Sephardi Y-chromosomes, appears to be one of the major founding lineages of the Palestinian and the Jewish population."Paragroup E-M35 * and haplogroup J-12f2a* fit the criteria for major AJ founding lineages because they are widespread both in AJ populations and in Near Eastern populations, and occur at much lower frequencies in European non-Jewish populations."
The following table only includes sample populations with more than 1% E-M215 men with all known subclades as of June 2015. It contains the E-V1515 clade defined by Trombetta et al. 2015, and all the E1b1b subclades distributed below the Sahara (E-V42, E-M293, E-V92, E-V6), which were identified as E-M35 basal clades in a former phylogeny.
| Population | N | Region | Language | Total | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-M215 | E-V2009 | E-M78* | E-V1477 | E-V1083* | E-V13 | E-V22 | E-V12* | E-V32 | E-V259 | E-V65 | E-V257* | E-M81 | E-M123* | E-M34 | E-V1515* | E-V1486* | E-V2881* | E-V1792 | E-V92 | E-M293* | E-V3065 | E-V42 | E-V1785* | E-V6 | E-V16 | ||||
| Northern Africa | |||||||||||||||||||||||||||||
| Zriba Arabs | 31 | Tunisia | AA/Semitic | 100 | |||||||||||||||||||||||||
| Reguibate Arabs | 20 | Algeria | AA/Semitic | 80 | |||||||||||||||||||||||||
| Kairouan Arabs | 49 | Tunisia | AA/Semitic | 89.6 | |||||||||||||||||||||||||
| Jerba Arabs | 47 | Tunisia | AA/Semitic | 93.6 | |||||||||||||||||||||||||
| Moroccan Arabs | 55 | Morocco | AA/Semitic | 15.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.3 | 0.0 | 0.0 | 0.0 | 32.7 | 0.0 | 30.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Asni Berbers | 54 | Morocco | AA/Berber | 85.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.9 | 79.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bouhria Berbers | 67 | Morocco | AA/Berber | 79.1 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 77.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Middle Atlas Berbers | 69 | Morocco | AA/Berber | 81.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.1 | 0.0 | 71.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Marrakech Berbers | 27 | Morocco | AA/Berber | 92.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 3.7 | 0.0 | 0.0 | 0.0 | 3.7 | 77.8 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Souss Berbers | 34 | Morocco | AA/Berber | 79.4 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 76.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ouarzazate Berbers | 31 | Morocco | AA/Berber | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Mozabite Berbers | 67 | Algeria | AA/Berber | 89.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 86.6 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Tunisian Jews | 10 | Tunisia | AA/Semitic | 20.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Libyan Arabs | 10 | Libya | AA/Semitic | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Libyan Jews | 23 | Libya | AA/Semitic | 26.1 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 17.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Northern Egyptians | 49 | Egypt | AA/Semitic | 20.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.0 | 16.3 | 4.1 | 2.0 | 0.0 | 0.0 | 0.0 | 4.1 | 4.1 | 10.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Egyptian Berbers from Siwa | 93 | Egypt | AA/Semitic | 18.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 4.3 | 2.2 | 1.1 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.5 | 0.0 |
| Egyptians from Baharia | 41 | Egypt | AA/Semitic | 56.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 22.0 | 14.6 | 0.0 | 0.0 | 2.4 | 7.3 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 |
| Egyptians from Gurna Oasis | 34 | Egypt | AA/Semitic | 17.6 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 8.8 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southern Egyptians | 47 | Egypt | AA/Semitic | 78.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 74.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Western/Central Africa | |||||||||||||||||||||||||||||
| Mandenka | 16 | Senegal | NC/Mande | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Tuareg | 22 | Niger | AA/Berber | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Daba | 29 | Cameroon (North) | AA/Chadic | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Guidar | 9 | Cameroon (North) | AA/Chadic | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Mandara | 82 | Cameroon (North) | AA/Chadic | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Shuwa Arabs | 5 | Cameroon (North) | AA/Semitic | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Fulbe from Cameroon | 76 | Cameroon (North) | NC/Atlantic | 1.3 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Moundang | 21 | Cameroon (North) | NC/Adamawa | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Eastern Africa | |||||||||||||||||||||||||||||
| Tigre | 5 | Eritrea | AA/Semitic | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 |
| Nara | 15 | Eritrea | NS/Sudanic | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 13.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 0.0 | 6.7 | 0.0 |
| Cunama | 20 | Eritrea | NS/Cunama | 65.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.0 | 10.0 | 5.0 | 0.0 |
| Saho | 94 | Eritrea | AA/Cushitic | 98.9 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 88.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.5 | 0.0 |
| Tigrai | 32 | Eritrea/Ethiopia | AA/Semitic | 71.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 21.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 31.3 | 6.3 | 0.0 |
| Afar | 25 | Djibouti | AA/Cushitic | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 56.0 | 0.0 |
| Somali | 40 | Djibouti | AA/Cushitic | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ethiopian Jews | 22 | Ethiopia | AA/Cushitic | 31.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 |
| Amhara | 82 | Ethiopia | AA/Semitic | 45.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 11.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.4 | 0.0 | 0.0 | 2.4 | 0.0 | 1.2 | 0.0 | 0.0 | 1.2 | 0.0 | 8.5 | 4.9 |
| Oromo | 62 | Ethiopia | AA/Cushitic | 53.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 17.7 | 0.0 | 0.0 | 1.6 | 0.0 | 3.2 | 0.0 | 1.6 | 1.6 |
| Wolayta | 12 | Ethiopia | AA/Omotic | 58.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Somali | 12 | Ethiopia | AA/Cushitic | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Gurage | 7 | Ethiopia | AA/Semitic | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 28.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 14.3 | 0.0 |
| Somali | 5 | Somalia | AA/Cushitic | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 80.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Turkana | 6 | Kenya | NS/Sudanic | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Borana | 9 | Kenya | AA/Cushitic | 77.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Somali | 6 | Kenya | AA/Cushitic | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Nilotic Western Kenya | 11 | Kenya | NS/Sudanic | 45.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 |
| Luhya | 51 | Kenya | NC/Bantu | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Other Bantu | 17 | Kenya | NC/Bantu | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Kikuyu | 9 | Kenya | NC/Bantu | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Maasai | 45 | Kenya | NS/Sudanic | 37.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 24.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Tutsi | 9 | Burundi | NC/Bantu | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southern Africa | |||||||||||||||||||||||||||||
| !Kung | 64 | Angola | KS | 10.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 |
| Khwe | 26 | Namibia | KS | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bantu | 8 | South Africa | NC/Bantu | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Europe | |||||||||||||||||||||||||||||
| Northern Portuguese | 50 | Portugal | IE | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southern Portuguese | 49 | Portugal | IE | 16.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Pasiegos from Cantabria | 56 | Spain | IE | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.8 | 41.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Asturians | 90 | Spain | IE | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 4.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southern Spaniards | 62 | Spain | IE | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.2 | 0.0 | 0.0 | 0.0 | 0.0 | 1.6 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Spanish Basques | 55 | Spain | Basque | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| French | 85 | France | IE | 8.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| French Basques | 16 | France | Basque | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Corsicans | 140 | France | IE | 6.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 | 0.0 | 0.0 | 1.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Danish | 35 | Denmark | IE | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Germans | 77 | Germany | IE | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Northern Italians | 80 | Italy | IE | 11.3 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.3 | 0.0 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Central Italians | 356 | Italy | IE | 12.9 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 2.0 | 0.3 | 0.0 | 0.0 | 0.3 | 0.3 | 0.8 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southern Italians | 141 | Italy | IE | 15.6 | 0.7 | 0.0 | 0.0 | 0.0 | 8.5 | 1.4 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.4 | 0.0 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Sicilians | 153 | Italy | IE | 20.3 | 0.0 | 0.0 | 0.0 | 0.0 | 7.2 | 4.6 | 0.7 | 0.0 | 0.0 | 0.7 | 0.0 | 0.7 | 0.0 | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Sardinians | 374 | Italy | IE | 8.3 | 0.8 | 0.0 | 0.0 | 0.3 | 1.1 | 0.8 | 0.3 | 0.0 | 0.0 | 1.1 | 0.3 | 0.3 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Polish | 40 | Poland | IE | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Slovenians | 104 | Slovenia | IE | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Estonians | 74 | Estonia | U | 5.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Hungarians | 106 | Hungary | U | 10.4 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Romanians | 30 | Romania | IE | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Macedonians | 99 | Macedonia | IE | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Continental Greeks | 32 | Greece | IE | 28.1 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bulgarians | 112 | Bulgaria | IE | 22.3 | 0.0 | 0.0 | 0.0 | 0.0 | 21.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Sephardic Bulgarians | 20 | Bulgaria | IE | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Albanians | 21 | Albania | IE | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Near East | |||||||||||||||||||||||||||||
| Sephardic Turkish | 19 | Turkey | A | 10.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 0.0 | 5.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Istanbul Turkish | 35 | Turkey | A | 17.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 5.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.7 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southwestern Turkish | 40 | Turkey | A | 7.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Northeastern Turkish | 41 | Turkey | A | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Central Anatolian | 61 | Turkey | A | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Southeastern Turkish | 24 | Turkey | A | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Erzurum Turkish | 25 | Turkey | A | 12.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Turkish Cypriots | 46 | Turkey | A | 23.9 | 0.0 | 0.0 | 0.0 | 0.0 | 10.9 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.7 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bedouins | 28 | Israel | AA/Semitic | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 7.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Druze Arabs | 28 | Israel | AA/Semitic | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 10.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Palestinians | 29 | Israel | AA/Semitic | 13.8 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 6.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Emiratis | 41 | United Arab Emirates | AA/Semitic | 7.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Omanites | 13 | Oman | AA/Semitic | 15.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Yemenites | 94 | Yemen | AA/Semitic | 14.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 3.2 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 7.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 |
Exceptional cases of men who are M215 positive but M35 negative ("E-M215*") have been discovered so far in two Amharas of Ethiopia and one Yemeni. At least some of these men, perhaps all, are known since early 2011 to be in a rare sibling clade to E-M35, known as E-V16 or E-M281. The discovery of M281 was announced by , who found it in two Ethiopian Oromo. found 5 more Ethiopian individuals and an equivalent SNP to M281, V16. It was in the 2011 paper that the family tree position (M215+/M35-) was discovered as described above. The E-M215 derivative, E-M35 is defined by the M35 SNP. 1 Turkmen individual from Jawzjan with a subclade defining mutation is referred to as E-M35*. As of June 2015, there is an increasingly complex tree structure which divides most men in E-M35 into two branches: E-V68 and E-Z827.
The most frequently described subclades are E-M78, a part of E-V68, and E-M81, which is a branch of E-Z827. These two subclades represent the largest proportion of the modern E-M215 population. E-M78 is found over most of the range where E-M215 is found excluding Southern Africa. E-M81 is found mainly in North Africa. E-M123 is less common but widely scattered, with significant populations in specific parts of the Horn of Africa, the Levant, Arabia, Iberia, and Anatolia. A new clade (E-V1515) was defined by Trombetta et al. 2015, which originated about 12 kya (95% CI 8.6–16.4) in eastern Africa where it is currently mainly distributed. This clade includes the E-V42, E-M293, E-V92 and E-V6 subclades, which were identified as E-M35 basal clades in a previous phylogeny.
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257. Both contain a lineage which has been frequently observed in Africa (E-M78 and E-M81, respectively) and a group of undifferentiated chromosomes that are mostly found in southern Europe. An expansion of E-M35 carriers, possibly from the Middle East as proposed by other authors, and split into two branches separated by the geographic barrier of the Mediterranean Sea, would explain this geographic pattern. However, the absence of E-V68* and E-V257* in the Middle East makes a maritime spread between northern Africa and southern Europe a more plausible hypothesis.
.png)
TMRCA of the major nodes in E-M35
| TMRCA (kya) | Trombetta 2015 | YFull |
|---|---|---|
| E-M215 | 39 | 35,4 |
| *E-M35 | 25 | 23,9 |
| **E-V68 | 20 | 20 |
| ***E-M78 | 15 | 13 |
| **E-Z827 | ? | 23,6 |
| ***E-V257/L19 | ? | 13,9 |
| ****E-M81 | ? | 2,7 |
| ***E-Z830 | 20 | 19 |
| ****E-M34 | ? | 15 |
| ****E-V1515 | 19 | ? |
Subclades
E-V68 (E1b1b1a)
Main article: Haplogroup E-V68
E-V68, is dominated by its longer-known subclade E-M78. Three "E-V68*" individuals who are in E-V68 but not E-M78 have been reported in Sardinia, by , when announcing the discovery of V68. The authors noted that because E-V68* was not found in the Middle Eastern samples, this appears to be evidence of maritime migration from Africa to southwestern Europe. E-M78 is a commonly occurring subclade, widely distributed in North Africa, the Horn of Africa, West Asia, (the Middle East and Near East) "up to Southern Asia", The European distribution has a frequency peak centered in parts of the Balkans (up to almost 50% in some areas) and Sicily, and declining frequencies evident toward western, central, and northeastern Europe.
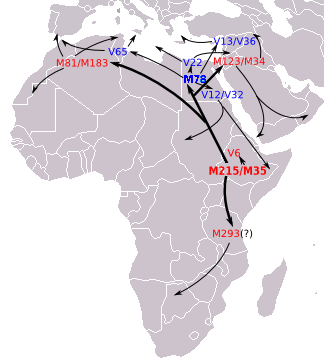
Based on genetic STR variance data, suggests that E-M78 originated in the region of Egypt and Libya. use the term Northeastern Africa to refer to Egypt and Libya, as shown in Table 1 of the study. Prior to , East Africa as a possible place of origin of E-M78, based upon Ethiopian testing. This was because of the high frequency and diversity of E-M78 lineages in the region of Ethiopia. However, were able to study more data, including populations from North Africa who were not represented in the study, and found evidence that the E-M78 lineages which make up a significant proportion of some populations in that region, were relatively young branches (see E-V32 below). They therefore concluded that "Northeast Africa" was the likely place of origin of E-M78 based on "the peripheral geographic distribution of the most derived subhaplogroups with respect to northeastern Africa, as well as the results of quantitative analysis of UEP and microsatellite diversity". So according to E-M35, the parent clade of E-M78, originated in East Africa, subsequently spread to Northeast Africa, and then there was a "back migration" of E-M215 chromosomes that had acquired the E-M78 mutation. therefore note this as evidence for "a corridor for bidirectional migrations" between Northeast Africa (Egypt and Libya in their data) on the one hand and East Africa on the other. The authors believe there were "at least 2 episodes between 23.9–17.3 ky and 18.0–5.9 ky ago". about 18,600 years ago (17,300 – 20,000 years ago). use two calculation methods for estimating the age of E-M78 which give very different results. For the main 18,600 years ago, the ASD_method is used, while for a second "Ρ_method", used as a check, gives 13.7kya with a Standard_deviation of 2.3kya, but the difference between the two methods is only large for the age estimation of E-M78, not its subclades. The authors state that the big difference is "attributable to the relevant departure from a star-like structure because of repeated founder effects" describe Egypt as "a hub for the distribution of the various geographically localized M78-related subclades" and, based on archaeological data, they propose that the point of origin of E-M78 (as opposed to later dispersal from Egypt) may have been in a refugium which "existed on the border of present-day Sudan and Egypt, near Lake Nubia, until the onset of a humid phase around 8500 BC. The northward-moving rainfall belts during this period could have also spurred a rapid migration of Mesolithic foragers northwards in Africa, the Levant and ultimately onward to Asia Minor and Europe, where they each eventually differentiated into their regionally distinctive branches". Towards the south, also explain evidence that some subclades of E-M78, specifically E-V12 and E-V22, "might have been brought to Sudan from North Africa after the progressive desertification of the Sahara around 6,000–8,000 years ago". And similarly, propose that E-M78 in Ethiopia, Somalia and surrounding areas, back-migrated to this region from the direction of Egypt after acquiring the E-M78 mutation.
Recently, E-M78 was dated by Trombetta et al. 2015. between 20,300 and 14,800 years ago.
Subclades of E-M78
Listed here are the main subclades of M78 as of June 2015. Within the E-M78 subclade, Trombetta et al. 2015 allocated most of the former E-M78* chromosomes to three new distinct branches: E-V1083*, E-V1477 and E-V259. The first is a paragroup sister to clades E-V22 and E-V13. The mutation V1477 defines a new basal branch that has been observed only in one northern African sample. Finally, a sister clade of E-V12 defined by V264 includes E-V65 and V259, a new lineage distributed in central Africa.
- E-M78 (E1b1b1a1) North Africa, Horn of Africa, West Asia, Europe (formerly "E1b1b1a").
- E-M78* Found in Morocco, southern Portugal, southern Spain and Iran (Tehran and Semnan provinces).
- E-V1477 Found in Tunisian Jews.
- E-V1083
- E-V1083* Found only in Eritrea (1.1%) and Sardinia (0.3%).
- E-V13 This is the most common subclade of E-M215 found in Europe. It is especially common in the Balkans.
- E-V22. Concentrated in Northeast Africa and the Near East. Peaks among the Saho.
- E-V1129
- E-V12. Found in Egypt, Sudan, and Chad other places. Has an important subclade
- E-V12* Most common lineage among Southern Egyptians (74.5%).
- E-V32. Very common among Somalis, Tigre and Oromos.
- E-V264
- E-V259 Found in North Cameroon.
- E-V65 Associated with North Africa, but also found in Sicily and also found in continental Italy.
- E-V12. Found in Egypt, Sudan, and Chad other places. Has an important subclade
- E-M521 Not mentioned by Trombetta et al.2015. Found in two individuals in Greece by and in one individual from the Eastern Alpine region of Italy by Coia et al. (2013)
E-Z827 (E1b1b1b)
Main article: Haplogroup E-Z827
In human genetics, E-Z827, is the name of a major human Y-chromosome DNA haplogroup abundantly found in North Africa, particularly the Maghreb, and to a lesser extent in Horn of Africa, the Near East and Europe.
E-L19 (E1b1b1b1)

To date, only two immediate downstream subclades of E-L19 have been recognized: E-PF2431 and E-M81.
'''E-M81'''
E-M81 is the dominant subclade of E-L19, accounting for more than 99% of its carriers. It has three main branches: E-M165, carried exclusively by a German and an Israeli; E-Y596059, carried by a Syrian from Damascus and an Iraqi Al Anbar; and E-CTS4236, carried exclusively by a Syrian from Quneitra, nearly all branches of E-M81 today derives from E-CTS4236. It is thought to have originated in the Near East, roughly 13,000 years ago, with subsequent expansions into Europe and North Africa occurring either about 1,000 years ago via Arab populations or around 3,000 years ago through Phoenician movements.This distribution has been usually explained as a consequence of a westward expansion from the Near East and this event probably occurred in recent times, possibly about 2 kya [22]. We calculated the forensic indexes including only the E-M81 subjects and we could observe an even more enhanced clinal pattern (Moroccan DC = 0.87; Libyan DC = 0.96; the values for the Egyptian populations cannot be considered informative because our collection included only four E-M81 chromosomes from this country) (Supplementary Table 12).This haplogroup has been identified in a variety of historically significant populations, including the ancient Phoenicians from Malaka and Selinunte, Medieval Arab communities from Al-Andalus and the Emirate of Sicily, as well as among the indigenous population of the Canary Islands.
| ID | Language | Culture | Country | Location | Date | Study | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I19190 | Semitic | Phoenician | Italy | Selinunte | 700-400BC | Ringbauer at al. 2025 | |||||||||||||||||||||||||||||||||
| I3682 | Semitic | Punico-Roman | Spain | Lucena | 300-400AD | title=Disparate demographic impacts of the Roman Colonization and the Migration Period in the Iberian Peninsula | date=2024-01-01 | biorxiv=10.1101/2024.09.23.614606 | language=en | first1=Pablo | last1=Carrión | first2=Iñigo | last2=Olalde | first3=Juan Manuel | last3=Jiménez-Arenas | first4=Neus | last4=Coromina | first5=David | last5=Vivó | first6=Josep María | last6=Vergès | first7=Ana | last7=Costa | first8=Daniel | last8=Botella | first9=Macarena | last9=Bustamante-Álvarez | first10=Javier | last10=Heras-Mora | first11=Ricardo | last11=Ortega-Ruiz | first12=Celia | last12=Chaves | first13=Maite Iris | last13=García-Collado | first14=Juan Antonio | last14=Quirós-Castillo | first15=Jordi | last15=Roig}} |
| I21367 | Semitic | Punico-Roman | Spain | Malaga | 416-700AD | Carrion et al. 2024 | |||||||||||||||||||||||||||||||||
| GUN012 | Unknown | Guanche | Spain | Tenerife | 593-660 AD | title=Genomic Analyses of Pre-European Conquest Human Remains from the Canary Islands Reveal Close Affinity to Modern North Africans | journal=Current Biology: CB | date=2017-11-06 | issn=1879-0445 | pmid=29107554 | pages=3396–3402.e5 | volume=27 | issue=21 | doi=10.1016/j.cub.2017.09.059 | first1=Ricardo | last1=Rodríguez-Varela | first2=Torsten | last2=Günther | first3=Maja | last3=Krzewińska | first4=Jan | last4=Storå | first5=Thomas H. | last5=Gillingwater | first6=Malcolm | last6=MacCallum | first7=Juan Luis | last7=Arsuaga | first8=Keith | last8=Dobney | first9=Cristina | last9=Valdiosera | first10=Mattias | last10=Jakobsson | first11=Anders | last11=Götherström | first12=Linus | last12=Girdland-Flink | bibcode=2017CBio...27E3396R }} |
| SP7089 | Semitic | Andalusian | France | Narbonne | 637-765 AD | Gleize et al. 2016 | |||||||||||||||||||||||||||||||||
| SP7080 | Semitic | Andalusian | France | Narbonne | 684-761 AD | Gleize et al. 2016 | |||||||||||||||||||||||||||||||||
| 78 | Semitic | Andalusian | Spain | Pamplona | 680-780 AD | de Miguel bánez 2016 | |||||||||||||||||||||||||||||||||
| 107 | Semitic | Andalusian | Spain | Pamplona | 680-780 AD | de Miguel Ibanez 2016 | |||||||||||||||||||||||||||||||||
| GUN011 | Unknown | Guanche | Spain | Tenerife | 704-887 AD | Rodriguez-Varela et al. 2017 | |||||||||||||||||||||||||||||||||
| GOG23 | Semitic | Andalusian | Spain | Vall Uixo | 706-888 AD | Oteo-Garcia et al. 2024 | |||||||||||||||||||||||||||||||||
| CAN02 | Unknown | Guanche | Spain | La Gomera | 774-994 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| ST2933 | Indo-European | Flemish | Belgium | Sint Truiden | 1020-1160 AD | Beneker et al. 2025 | |||||||||||||||||||||||||||||||||
| CAN048 | Unknown | Guanche | Spain | Tenerife | 1028-1162 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| GUN002 | Unknown | Guanche | Spain | Tenerife | 1031-1159 AD | Rodriguez-Varela et al. 2017 | |||||||||||||||||||||||||||||||||
| SGBN3 | Semitic | Andalusian | Italy | Segesta | 1050-1215 AD | Monnereau et al. 2024 | |||||||||||||||||||||||||||||||||
| SGBN1 | Semitic | Andalusian | Italy | Segesta | 1050-1220 AD | Monnereau et al. 2024 | |||||||||||||||||||||||||||||||||
| 112644 | Semitic | Andalusian | Spain | Sagunto | 1100-1300 AD | Olalde et al. 2019 | |||||||||||||||||||||||||||||||||
| 112649 | Semitic | Andalusian | Spain | Sagunto | 1100-1300 AD | Olalde et al. 2019 | |||||||||||||||||||||||||||||||||
| CAN012 | Unknown | Guanche | Spain | Gran Canaria | 1169-1265 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| PT23221 | Semitic | Andalusian | Portugal | Santarem | 1200-1300 AD | Roca-Rada et al. 2024 | |||||||||||||||||||||||||||||||||
| CAN023 | Unknown | Guanche | Spain | Gran Canaria | 1221-1278 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN041 | Unknown | Guanche | Spain | Tenerife | 1228-1297 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN001 | Unknown | Guanche | Spain | Tenerife | 1272-1388 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN030 | Unknown | Guanche | Spain | La Gomera | 1288-1396 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN025 | Unknown | Guanche | Spain | Gran Canaria | 1303-1410 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN043 | Unknown | Guanche | Spain | Gran Canaria | 1322-1437 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN042 | Unknown | Guanche | Spain | Tenerife | 1327-1440 AD | Serrano et al. 2023 | |||||||||||||||||||||||||||||||||
| CAN045 | Unknown | Guanche | Spain | Gran Canaria | 1444-1625 AD | Serrano et al,.2023 | |||||||||||||||||||||||||||||||||
| ELW036 | Indo-European | German | Germany | Ellwangen | 1486-1627 AD | Immel et al. 2021 | |||||||||||||||||||||||||||||||||
| GOG59 | Indo-European | Andalusian | Spain | Vall Uixo | 1501-1600 AD | Oteo-Garcia et al. 2024 | |||||||||||||||||||||||||||||||||
| PT22190 | Indo-European | Portuguese | Spain | Aveiro | 1701-1800 AD | Roca-Rada et al. 2024 |
'''E-PF2431'''
E-PF2431 is a minor subclade of E-L19 mostly found in Europe, with smaller but notable frequencies in the Middle East/North Africa and sub-Saharan Africa. This haplogroup has been identified among Phoenicians from Motya, ancient Romans from Pompeii, and Medieval Arabs.
E-Z830 (E1b1b1b2)
This is a recently discovered subclade which has not yet been included in most haplogroup trees, E-Z830 includes the confirmed subclades of E-M123, E-V1515 (E-M293, E-V42, E-V6, E-V92), and E-Z830*, and is a sibling clade to E-L19. Currently, the E-M35 phylogeny project] recognizes four distinct clusters of Z830* carriers, two of which are exclusively Jewish in origin. The remaining two are significantly smaller, and include scattered individuals in Germany, Spain, Latin America, Egypt, and Ethiopia.
E-M123
Main article: Haplogroup E-M123
E-M123 is mostly known for its major subclade E-M34, which dominates this clade.As of 11 November 2008 for example, the E-M35 phylogeny project had records of four E-M123* tests, compared to 93 test results with E-M34.
E-V1515
A new clade (E-V1515) was defined by Trombetta et al. 2015, which originated about 12 kya (95% CI 8.6–16.4) in eastern Africa where it is currently mainly distributed. This clade includes all the sub-Saharan haplogroups (E-V42, E-M293, E-V92, E-V6) reported as E-M35 basal clades in a previous phylogeny.
E-M293
E-M293 is a subclade of E-V1515. It was first identified by ISOGG as the second clade within E-Z830. It was discovered before E-Z830, being announced in , which associated it with the spread of pastoralism from East Africa into Southern Africa. So far high levels have been found in specific ethnic groups in Tanzania and Southern Africa. Highest were the Datooga (43%), Khwe (Kxoe) (31%), Burunge (28%), and Sandawe peoples (24%). Henn (2008) in their study also found two Bantu-speaking Kenyan males with the M293 mutation.
Other E-M215 subclades are rare in Southern Africa. The authors state "Without information about M293 in the Maasai, Hema, and other populations in Kenya, Sudan, and Ethiopia, we cannot pinpoint the precise geographic source of M293 with greater confidence. However, the available evidence points to present-day Tanzania as an early and important geographic locus of M293 evolution.". They also say that "M293 is only found in sub-Saharan Africa, indicating a separate phylogenetic history for M35.1 * (former) samples further north".
E-P72 appears in Karafet (2008). announced that this is a subclade of E-M293.
E-V42
announced the discovery of E-V42 in two Beta Israel persons. It was suggested that it may be restricted to the region around Ethiopia. However, further testing by commercial DNA testing companies confirmed many positive results for this subclade in Saudi Arabia, Kuwait and one person in Portugal who has a root from Arabia.
E-V6
The E-V6 subclade of E-V1515 is defined by V6. identified a significant presence of these lineages in Ethiopia and also some in the neighboring Somalis. Among the Ethiopian and Somali samples, the highest were 14.7% among the Amhara and 16.7% among the Wolayta.
To the south, identified one V6+ man in a sample of 35 Datooga of Tanzania. And further to the north, identified another 6 men in a sample of 93 from the Siwa Oasis, which is a Berber population
E-V92
announced the discovery of E-V92 in two Amharas. Like E-V6 and E-V42 it possibly only exists in the area of Ethiopia.
Phylogenetics
Phylogenetic history
Main article: Conversion table for Y chromosome haplogroups
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) |
|---|---|---|---|
| (δ) | (ε) | (ζ) | (η) |
| E-P29 | 21 | III | 3A |
| E-M33 | 21 | III | 3A |
| E-M44 | 21 | III | 3A |
| E-M75 | 21 | III | 3A |
| E-M54 | 21 | III | 3A |
| E-P2 | 25 | III | 4 |
| E-M2 | 8 | III | 5 |
| E-M58 | 8 | III | 5 |
| E-M116.2 | 8 | III | 5 |
| E-M149 | 8 | III | 5 |
| E-M154 | 8 | III | 5 |
| E-M155 | 8 | III | 5 |
| E-M10 | 8 | III | 5 |
| E-M35 | 25 | III | 4 |
| E-M78 | 25 | III | 4 |
| E-M148 | 25 | III | 4 |
| E-M81 | 25 | III | 4 |
| E-M107 | 25 | III | 4 |
| E-M165 | 25 | III | 4 |
| E-M123 | 25 | III | 4 |
| E-M34 | 25 | III | 4 |
| E-M136 | 25 | III | 4 |
Research publications
The following research teams per their publications were represented in the creation of the YCC Tree.
- α and
- β
- γ
- δ
- ε
- ζ
- η
Phylogenetic trees
Cladogram with the main subclades:
The following phylogenetic tree is based on the YCC 2008 tree and subsequent published research as summarized by ISOGG. It includes all known subclades as of June 2015 (Trombetta et al. 2015)
- E-M215 (E1b1b)
- E-M215*. Rare or non-existent.
- E-M35 (E1b1b1)
- E-V68 (E1b1b1a)
- E-V2009. Found in individuals in Sardinia and Morocco.
- E-M78 (E1b1b1a1). North Africa, Horn of Africa, West Asia, Sicily. (Formerly "E1b1b1a".)
- E-M78*
- E-V1477. Found in Tunisian Jews.
- E-V1083.
- E-V1083*. Found only in Eritrea (1.1%) and Sardinia (0.3%).
- E-V13
- E-V22
- E-V1129
- E-V12
- E-V12*
- E-V32
- E-V264
- E-V259. Found in North Cameroon.
- E-V65
- E-CTS194
- E-V12
- E-Z827 (E1b1b1b)
- E-V257/L19 (L19, V257) – E1b1b1b1
- E-PF2431
- E-M81 (M81)
- E-PF2546
- E-PF2546*
- E-CTS12227
- E-MZ11
- E-MZ12
- E-MZ11
- E-A929
- E-Z5009
- E-Z5009*
- E-Z5010
- E-Z5013
- E-Z5013*
- E-A1152
- E-A2227
- E-A428
- E-MZ16
- E-PF6794
- E-PF6794*
- E-PF6789
- E-MZ21
- E-MZ23
- E-MZ80
- E-A930
- E-Z2198/E-MZ46
- E-A601
- E-L351
- E-Z5009
- E-PF2546
- E-Z830 (Z830) – E1b1b1b2
- E-M123 (M123)
- E-M34 (M34)
- E-M84 (M84)
- E-M136 (M136)
- E-M290 (M290)
- E-V23 (V23)
- E-L791 (L791,L792)
- E-M84 (M84)
- E-M34 (M34)
- E-V1515. E-V1515 and its subclades are mainly restricted to eastern Africa.
- E-V1515*
- E-V1486
- E-V1486*
- E-V2881
- E-V2881*
- E-V1792
- E-V92
- E-M293 (M293)
- E-M293*
- E-P72 (P72)
- E-V3065*
- E-V1700
- E-V42 (V42)
- E-V1785
- E-V1785*
- E-V6 (V6)
- E-M123 (M123)
- E-V257/L19 (L19, V257) – E1b1b1b1
- E-V16/E-M281 (E1b1b2). Rare. Found in individuals in Ethiopia, Yemen and Saudi Arabia.
- E-V68 (E1b1b1a)
Notes
References
Bibliography
- . Also at http://www.ucl.ac.uk/tcga/tcgapdf/Behar-AJHG-03.pdf and https://web.archive.org/web/20090304100321/http://www.familytreedna.com/pdf/400971.pdf
- also at http://www.ucl.ac.uk/tcga/tcgapdf/capelli-CB-03.pdf
- Also see Supplementary Data.
- . See comment on Dienekes blog, comment on the Spitoon blog and public release .
- . Published online April 2, 2008. See also Supplementary Material.
- . (Also see Errata)
- .
- Pontikos D. "Phylogeographic refinement of haplogroup E" http://dienekes.blogspot.ru/2015/07/phylogeographic-refinement-of.html
- . Published online 9 March 2005
- .
- .
Sources for conversion tables
References
- "E-M35 YTree".
- {{Harvcoltxt. Cruciani. La Fratta. Santolamazza. Sellitto. 2004
- {{Harvcoltxt. Cruciani. La Fratta. Trombetta. Santolamazza. 2007
- {{Harvcoltxt. Henn. Gignoux. Lin. Oefner. 2008
- {{Harvcoltxt. Hassan. Underhill. Cavalli-Sforza. Ibrahim. 2008
- (July 2015). "Phylogeographic Refinement and Large Scale Genotyping of Human Y Chromosome Haplogroup E Provide New Insights into the Dispersal of Early Pastoralists in the African Continent". Genome Biology and Evolution.
- [http://www.nature.com/ncomms/2015/150519/ncomms8152/fig_tab/ncomms8152_T1.html Large-scale recent expansion of European patrilineages shown by population resequencing], Chiara Batini et al, ''nature.com'', 2015
- {{Harvcoltxt. Lacan. Keyser. Ricaut. Brucato. 2011
- "Molecular genetic investigation of the Neolithic population history in the western Carpathian Basin".
- (4 May 2018). "Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations". Science.
- (February 2022). "Placing Ancient DNA Sequences into Reference Phylogenies". Molecular Biology and Evolution.
- (2022). "Science". Science.
- (2024-04-26). "Palaeogenomic insights into the origins of early settlers on the island of Cyprus". Scientific Reports.
- (2018). "PNAS". Proceedings of the National Academy of Sciences of the United States of America.
- (2020-05-28). "The Genomic History of the Bronze Age Southern Levant". Cell.
- (2020-05-28). "Genomic History of Neolithic to Bronze Age Anatolia, Northern Levant, and Southern Caucasus". Cell.
- (July 2025). "Punic people were genetically diverse with almost no Levantine ancestors". Nature.
- (2019-03-15). "The genomic history of the Iberian Peninsula over the past 8000 years". Science.
- (August 2025). "Whole-genome ancestry of an Old Kingdom Egyptian". Nature.
- (2017). "Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods". Nature Communications.
- {{Harvcoltxt. Behar. Thomas. Skorecki. Hammer. 2003
- {{Harvcoltxt. Semino. Magri. Benuzzi. Lin. 2004
- {{Harvcoltxt. Behar. Garrigan. Kaplan. Mobasher. 2004
- {{Harvcoltxt. Shen. Lavi. Kivisild. Chou. 2004
- {{Harvcoltxt. Adams. Bosch. Balaresque. Ballereau. 2008
- {{Harvcoltxt. Nebel. Filon. Brinkmann. Majumder. 2001
- Cadenas. Zhivotovsky. Cavalli-Sforza. Underhill. 2007
- {{harvp. Trombetta. Cruciani. Sellitto. Scozzari. 2011
- (Figure S.7) J D Cristofaro et al., 2013, "Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge", http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0076748
- Cruciani. La Fratta. Torroni. Underhill. 2006
- Peričic. Lauc. Klarić. Rootsi. 2005
- "E-M165 YTree".
- "E-CTS4236 YTree".
- (August 2004). "A predominantly neolithic origin for Y-chromosomal DNA variation in North Africa". American Journal of Human Genetics.
- (November 2009). "Geographical structure of the Y-chromosomal genetic landscape of the Levant: a coastal-inland contrast". Annals of Human Genetics.
- (2017-01-01). "Neolithization of North Africa involved the migration of people from both the Levant and Europe".
- Rubio Araúna, Lara. (2017-12-14). "Genetic structure of North African human populations : A complex history of admixture".
- (2019-01-01). "Rapidly mutating Y-STRs in rapidly expanding populations: Discrimination power of the Yfiler Plus multiplex in northern Africa". Forensic Science International. Genetics.
- "The Prehistory of Language from the Perspective of the Y-Chromosome".
- Solé Morata, Neus. (2017-12-15). "Inferring recent human population history from a Y chromosome perspective". Universitat Pompeu Fabra.
- Penninx, Wim. "The male lines of the Maghreb: Phoenicians, Carthage, Muslim conquest and Berbers". Independent Publishing..
- (2019-01-01). "Rapidly mutating Y-STRs in rapidly expanding populations: Discrimination power of the Yfiler Plus multiplex in northern Africa". Forensic Science International: Genetics.
- (2024-07-24). "Multi-proxy bioarchaeological analysis of skeletal remains shows genetic discontinuity in a Medieval Sicilian community". Royal Society Open Science.
- (2023-08-15). "The genomic history of the indigenous people of the Canary Islands". Nature Communications.
- (July 2025). "Punic people were genetically diverse with almost no Levantine ancestors". Nature.
- (2024-01-01). "Disparate demographic impacts of the Roman Colonization and the Migration Period in the Iberian Peninsula".
- (2017-11-06). "Genomic Analyses of Pre-European Conquest Human Remains from the Canary Islands Reveal Close Affinity to Modern North Africans". Current Biology: CB.
- (2016-02-24). "Early Medieval Muslim Graves in France: First Archaeological, Anthropological and Palaeogenomic Evidence". PLOS ONE.
- Cervantes, Biblioteca Virtual Miguel de. "La "maqbara" de Pamplona (s. VIII). Aportes de la osteoarqueología al conocimiento de la islamización en la Marca Superior / María Paz de Miguel Ibáñez ; dirigida por Mauro Severo Hernández Pérez, Sonia Gutiérrez Lloret {{!}} Biblioteca Virtual Miguel de Cervantes".
- (2024-01-01). "Medieval genomes from eastern Iberia illuminate the role of Morisco mass deportations in dismantling a long-standing genetic bridge with North Africa".
- (2023-08-15). "The genomic history of the indigenous people of the Canary Islands". Nature Communications.
- (2025-05-20). "Urbanization and genetic homogenization in the medieval Low Countries revealed through a ten-century paleogenomic study of the city of Sint-Truiden.". Springer Nature Link.
- (2024-07-24). "Multi-proxy bioarchaeological analysis of skeletal remains shows genetic discontinuity in a Medieval Sicilian community". Royal Society Open Science.
- (2019-03-15). "The genomic history of the Iberian Peninsula over the past 8000 years". Science.
- (2025-08-18). "The genetic history of Portugal over the past 5,000 years". Genome Biology.
- (2021-09-27). "Analysis of Genomic DNA from Medieval Plague Victims Suggests Long-Term Effect of Yersinia pestis on Human Immunity Genes". Molecular Biology and Evolution.
- (2025-04-28). "Medieval genomes from eastern Iberia illuminate the role of Morisco mass deportations in dismantling a long-standing genetic bridge with North Africa". Genome Biology.
- (July 2025). "Punic people were genetically diverse with almost no Levantine ancestors". Nature.
- (2024-11-18). "Ancient DNA challenges prevailing interpretations of the Pompeii plaster casts". Current Biology.
- (2024-09-23). "Disparate demographic impacts of the Roman Colonization and the Migration Period in the Iberian Peninsula".
- [http://www.haplozone.net/e3b/index.php/project/query]{{Dead link. (April 2019)
- "E-M35 Project Data".
- "E-M35 Project Data".
- "E-M35 Project Data".
- "E-M35 Project Data".
- "E-M35 Project Data".
- {{Harvcoltxt. ISOGG. 2011
- {{Harvcoltxt. Karafet. Mendez. Meilerman. Underhill. 2008
- {{Harvcoltxt. Y Chromosome Consortium "YCC". 2002
- ISOGG 2015
This article was imported from Wikipedia and is available under the Creative Commons Attribution-ShareAlike 4.0 License. Content has been adapted to SurfDoc format. Original contributors can be found on the article history page.
Ask Mako anything about Haplogroup E-M35 — get instant answers, deeper analysis, and related topics.
Research with MakoFree with your Surf account
Create a free account to save articles, ask Mako questions, and organize your research.
Sign up freeThis content may have been generated or modified by AI. CloudSurf Software LLC is not responsible for the accuracy, completeness, or reliability of AI-generated content. Always verify important information from primary sources.
Report