From Surf Wiki (app.surf) — the open knowledge base
Bacteriorhodopsin
Protein used by single-celled organisms

Protein used by single-celled organisms
| Field | Value |
|---|---|
| Name | Bacteriorhodopsin |
| Organism | Halobacterium salinarum |
| image | Halobacterium salinarum bacteriorhodopsin PDB 6RQP.png |
| caption | Halobacterium salinarum bacteriorhodopsin homotrimer viewed from the cytoplasm. Bound retinal shown in black. |
| Symbol | bop |
| UniProt | P02945 |
Bacteriorhodopsin (Bop) is a protein used by Archaea, most notably by Haloarchaea, a class of the Euryarchaeota. It acts as a proton pump; that is, it captures light energy and uses it to move protons across the membrane out of the cell. The resulting proton gradient is subsequently converted into chemical energy.
Function
| image-width = 220
| image-left = 0
| image-top = 0
Bacteriorhodopsin is a light-driven H+ ion transporter found in some Haloarchaea, most notably Halobacterium salinarum (formerly known as syn. H. halobium). The proton-motive force generated by the protein is used by ATP synthase to generate adenosine triphosphate (ATP). By expressing Bacteriorhodopsin, the archaea cells are able to synthesise ATP in the absence of a carbon source.
Structure
Bacteriorhodopsin is a 27* *kDa integral membrane protein usually found in two-dimensional crystalline patches known as "purple membrane", which can occupy almost 50% of the surface area of the archaeal cell. The repeating element of the hexagonal lattice is composed of three identical protein chains, each rotated by 120 degrees relative to the others. Each monomer has seven transmembrane alpha helices and an extracellular-facing, two-stranded beta sheet.
Bacteriorhodopsin is synthesized as a protein precursor, known as bacterio-opsin, which is extensively modified after translation. The modifications are:
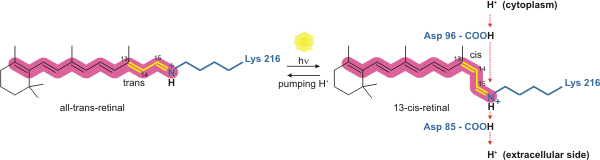
- Covalent conjugation of a retinal molecule to residue Lys216, via a Schiff base, to create the retinylidene chromophore.
- Cleavage of the signal peptide, the first 13 amino acids at the N-terminus, and the conversion of residue Gln14 to pyroglutamate
- Removal of residue Asp262 at the C-terminus
Spectral properties
Bacteriorhodopsin molecule is purple and is most efficient at absorbing green light (in the wavelength range 500-650 nm). In the native membrane, the protein has a maximum absorbance at 553 nm, however addition of detergent disrupts the trimeric form, leading a loss of exciton coupling between the chromophores, and the monomeric form consequently has an absorption maximum of 568 nm.
Bacteriorhodopsin has a broad excitation spectrum. For a detection wavelength between 700 and 800 nm, it has an appreciable detected emission for excitation wavelengths between 470 nm and 650 nm (with a peak at 570 nm). When pumped at 633 nm, the emission spectrum has appreciable intensity between 650 nm and 850 nm.
Mechanism
Photocycle overview
Bacteriorhodopsin is a light-driven proton pump. It is the retinal molecule that changes its isomerization state from all-trans to 13-cis when it absorbs a photon. The surrounding protein responds to the change in the chromophore shape, by undergoing an ordered sequence of conformational changes (collectively known as the photocycle). The conformational changes alter the pKa values of conserved amino acids in the core of the protein, including Asp85, Asp96 and the Schiff base N atom (Lys216). These sequential changes in acid dissociation constant, result in the transfer of one proton from the intracellular side to the extracellular side of the membrane for each photon absorbed by the chromophore.
The bacteriorhodopsin photocycle consists of nine distinct stages, starting from the ground or resting state, which is denoted 'bR'. The intermediates are identified by single letters and may be distinguished by their absorption spectra. The nine stages are: : bR + photon → K L M1 M2 M2' N N' O bR
Ground state + photon → K state → L state

Bacteriorhodopsin in the ground state absorbs a photon and the retinal changes isomerization from all-trans 15-anti to the strained 13-cis 15-anti in the K state. The isomerisation reaction is fast and occurs in less than 1 ps. The retinal adopts a less strained conformation to form the L intermediate.
L state → M1 state
Asp85 accepts a proton from the Schiff base N atom. In the M1 intermediate, neither the Schiff base nor Asp85 are charged.
M1 state → M2 state
The Schiff base rotates away from the extracellular side of the protein towards the cytoplasmic side, in preparation to accept a new proton.
M2 state → M2' state
A proton is released from Glu204 and Glu194 to the extracellular medium.
M2' state → N state
The retinal Schiff base accepts a proton from Asp96. In the N state, both Asp96 and the Schiff base are charged.
N state → N' state
Asp96 accepts a proton from the cytoplasmic side of the membrane and becomes uncharged.
N' state → O state
Retinal reisomerizes to the all-trans state.
O state → ground state
Asp85 transfers a proton to Glu194 and Glu204 on the extracellular face of the protein.
Homologs and other similar proteins
Bacteriorhodopsin belongs to the microbial rhodopsin family. Its homologs include the archaerhodopsins, the light-driven chloride pump halorhodopsin (for which the crystal structure is also known), and some directly light-activated channels such as channelrhodopsin.
Bacteriorhodopsin is similar to vertebrate rhodopsins, the pigments that sense light in the retina. Rhodopsins also contain retinal; however, the functions of rhodopsin and bacteriorhodopsin are different, and there is limited similarity in their amino acid sequences. Both rhodopsin and bacteriorhodopsin belong to the 7TM receptor family of proteins, but rhodopsin is a G protein-coupled receptor and bacteriorhodopsin is not. In the first use of electron crystallography to obtain an atomic-level protein structure, the structure of bacteriorhodopsin was resolved in 1990. It was then used as a template to build models of G protein-coupled receptors before crystallographic structures were also available for these proteins. It has been excessively studied on both mica and glass substrates using Atomic force microscopy and Femtosecond crystallography.
All other phototrophic systems in bacteria, algae, and plants use chlorophylls or bacteriochlorophylls rather than bacteriorhodopsin. These also produce a proton gradient, but in a quite different and more indirect way involving an electron transfer chain consisting of several other proteins. Furthermore, chlorophylls are aided in capturing light energy by other pigments known as "antennas"; these are not present in bacteriorhodopsin-based systems. It is possible that phototrophy independently evolved at least twice, once in bacteria and once in archaea.
Gallery
File:Bacteriorhodopsin subunit 1X0S.gif|link=File:Bacteriorhodopsin subunit 1X0S large.gif|Bacteriorhodopsin single monomer with retinal molecule between 7 vertical alpha helixes (PDB ID: 1X0S ). One more small helix is light blue, beta sheet yellow. File:Bacteriorhodopsin trimer 1X0S.png|link=File:Bacteriorhodopsin trimer 1X0S large.gif|Bacteriorhodopsin trimer with one retinal molecule in each subunit seen from the extracellular side EC (PDB ID: 1X0S )
Literature
References
- (2020). "Halobacteria". [[National Center for Biotechnology Information]].
- (2004). "Biochemistry". J. Wiley & Sons.
- "Bacteriorhodopsin: Pumping Ions".
- Nicholls DG. (1992). "Bioenergetics 2". Academic Press.
- Stryer, Lubert. (1995). "Biochemistry". WH Freeman and Company.
- (1998). "Lipid patches in membrane protein oligomers: Crystal structure of the bacteriorhodopsin-lipid complex". Proceedings of the National Academy of Sciences of the United States of America.
- (1997). "X-ray structure of bacteriorhodopsin at 2.5 Å from microcrystals grown in lipidic cubic phases". Science.
- (1999). "Structure of bacteriorhodopsin at 1.55 Å resolution". Journal of Molecular Biology.
- (1971). "Rhodopsin-like Protein from the Purple Membrane of Halobacterium halobium". Nature New Biology.
- (1982). "Reconstitution of the retinal proteins bacteriorhodopsin and halorhodopsin".
- (1981). "Site of attachment of retinal in bacteriorhodopsin". Proceedings of the National Academy of Sciences of the United States of America.
- (2021). "Detergent-free Lipodisq Nanoparticles Facilitate High-Resolution Mass Spectrometry of Folded Integral Membrane Proteins". Nano Letters.
- (2002). "Comparison of the Dynamics of the Primary Events of Bacteriorhodopsin in Its Trimeric and Monomeric States". Biophysical Journal.
- (2012). "The Exciton Origin of the Visible Circular Dichroism Spectrum of Bacteriorhodopsin". Journal of Physical Chemistry B.
- (2007). "On the excitation wavelength dependence of the fluorescence of bacteriorhodopsin". Chemical Physics Letters.
- (1995). "Fluorescence spectra of bacteriorhodopsin and the intermediates O and Q at room temperature". FEBS Lett..
- (September 2003). "Molecular dynamics simulation of bacteriorhodopsin's photoisomerization using ab initio forces for the excited chromophore". Biophysical Journal.
- (2014). "Microbial and animal rhodopsins: Structures, functions and molecular mechanisms". Chemical Reviews.
- (1998). "Existence of a proton transfer chain in bacteriorhodopsin: Participation of Glu-194 in the release of protons to the extracellular surface". Biochemistry.
- (1999). "The proton release group of bacteriorhodopsin controls the rate of the final step of its photocycle at low pH". Biochemistry.
- (2021). "Structures of the archaerhodopsin 3 transporter reveal that disordering of internal water networks underpins receptor sensitization". Nature Communications.
- Henderson R, Baldwin JM, Ceska TA, Zemlin F, Beckmann E, Downing KH. (1990). "Model for the structure of bacteriorhodopsin based on high-resolution electron cryo-microscopy". J Mol Biol.
- (2008). "Atomic force microscopy as a multifunctional molecular toolbox in nanobiotechnology". Nature Nanotechnology.
- (2010-02-14). "High-speed atomic force microscopy shows dynamic molecular processes in photoactivated bacteriorhodopsin". Nature Nanotechnology.
- (2016-12-23). "A three-dimensional movie of structural changes in bacteriorhodopsin". Science.
- (2005-03-28). "Crystal structure of the 13-cis isomer of bacteriorhodopsin.". RCSB Protein Data Bank (PDB).
- (2005). "Crystal structure of the 13-cis isomer of bacteriorhodopsin in the dark-adapted state.". J. Mol. Biol..
- Image created with [http://www.geneinfinity.org/rastop/ RasTop] (Molecular Visualization Software).
This article was imported from Wikipedia and is available under the Creative Commons Attribution-ShareAlike 4.0 License. Content has been adapted to SurfDoc format. Original contributors can be found on the article history page.
Ask Mako anything about Bacteriorhodopsin — get instant answers, deeper analysis, and related topics.
Research with MakoFree with your Surf account
Create a free account to save articles, ask Mako questions, and organize your research.
Sign up freeThis content may have been generated or modified by AI. CloudSurf Software LLC is not responsible for the accuracy, completeness, or reliability of AI-generated content. Always verify important information from primary sources.
Report